
-
Skin and subcutaneous tissue.
-
Muscles: a superficial layer, primarily
concerned with positioning and moving the limbs, and deeper layers
(“true back muscles”), specifically concerned with moving or
maintaining the position of the axial skeleton (posture). -
Vertebral column: the vertebrae, intervertebral (IV) discs, and associated ligaments (Fig. 4.1).
-
Ribs (in the thoracic region): particularly their posterior portions, medial to the angles of the ribs.
-
Spinal cord and meninges (membranes that cover the spinal cord).
-
Various segmental nerves and vessels.
back of the neck and the posterior and deep cervical muscles and
vertebrae are also described in this chapter. The scapulae, although
located in the back, are part of the appendicular skeleton and are
considered with the upper limb (Chapter 6).
(spine), which extends from the cranium (skull) to the apex of the
coccyx. The vertebral column forms the skeleton of the neck and back
and is the main part of the axial skeleton (i.e., the articulated bones of the cranium, vertebral column, ribs, and sternum) (Fig. 4.1D).
The adult vertebral column is 72–75 cm long, of which approximately one
quarter is formed by the IV discs, which separate and bind the
vertebrae together (Fig. 4.1D & E). The vertebral column:
-
Protects the spinal cord and spinal nerves.
-
Supports the weight of the body superior to the level of the pelvis.
-
Provides a partly rigid and flexible axis for the body and an extended base on which the head is placed and pivots.
-
Plays an important role in posture and locomotion (the movement from one place to another).
occur frequently in competitive sports and in industrial and automobile
accidents. In severe injuries, the examiner must be careful not to
cause further damage. For example, if an injured person complains of
back pain and is unable to move the limbs, the vertebral column may be
fractured. If the neck is flexed or the injured person sits up, the
spinal cord may be (further) injured. Improper handling of an injured
person can convert an unstable lesion without a neurological deficit
into one with a deficit that produces permanent disability.
33 vertebrae arranged in five regions: 7 cervical, 12 thoracic, 5
lumbar, 5 sacral, and 4 coccygeal (Fig. 4.1A–D).
Significant motion occurs only between the 25 superior vertebrae. Of
the 9 inferior vertebrae, the 5 sacral vertebrae are fused in adults to
form the sacrum and, after approximately age 30, the 4 coccygeal vertebrae fuse to form the coccyx. The lumbosacral angle occurs at the junction of, and is formed by, the long axes of the lumbar region of the vertebral column and the sacrum (Fig.4.1D).
The vertebrae gradually become larger as the vertebral column descends
to the sacrum and then become progressively smaller toward the apex of
the coccyx (Fig. 4.1A–D).
The change in size is related to the fact that successive vertebrae
bear increasing amounts of the body’s weight as the column descends.
The vertebrae reach maximum size immediately superior to the sacrum,
which transfers the weight to the pelvic girdle at the sacroiliac
joints.
which facilitate and control the vertebral column’s flexibility.
Although the movement between two adjacent vertebrae is small, in
aggregate the vertebrae and IV discs uniting them form a remarkably
flexible yet rigid column that protects the spinal cord they surround.
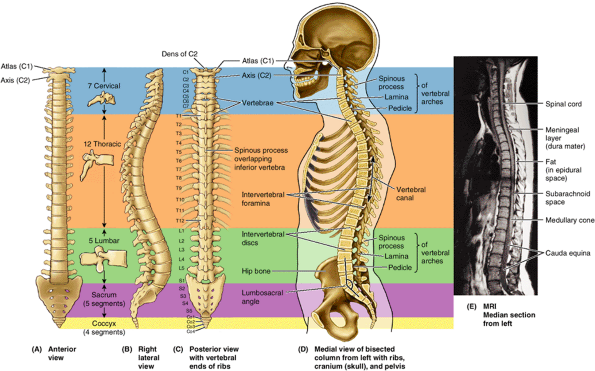 |
|
Figure 4.1. Vertebral column and vertebral canal, demonstrating its five regions. A. This anterior view shows the isolated vertebral column. B.
This right lateral view shows the isolated vertebral column. The isolated vertebrae are typical of each of the three mobile regions. Note the increase in size of the vertebrae as the column descends. C. This posterior view of the vertebral column includes the vertebral ends of ribs, representing the skeleton of the back. D. This medial view of the axial skeleton in situ demonstrates its regional curvatures and its relationship to the cranium (skull), thoracic cage, and hip bone. The continuous, weight-bearing column of vertebral bodies and IV discs forms the anterior wall of the vertebral canal. The lateral and posterior walls of the canal are formed by the series of vertebral arches. The IV foramina (seen also in part B) are openings in the lateral wall through which spinal nerves exit the vertebral canal. The posterior wall is formed by overlapping laminae and spinous processes, like shingles on a roof. E. This sagittal MRI study shows the primary contents of the vertebral canal. The medullary cone (L. conus medullaris) is the cone-shaped inferior end of the spinal cord, which typically ends at the L1–L2 level in adults. The dura mater, the external covering of the spinal cord (gray), is separated from the spinal cord by a fluid-filled space (black) and from the wall of the vertebral canal by fat (white) and thin-walled veins (not visible here). |
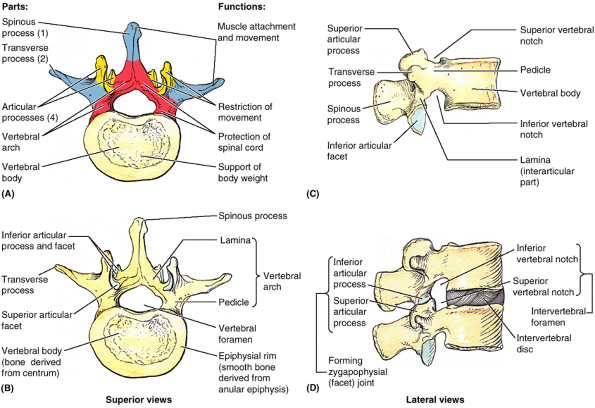 |
|
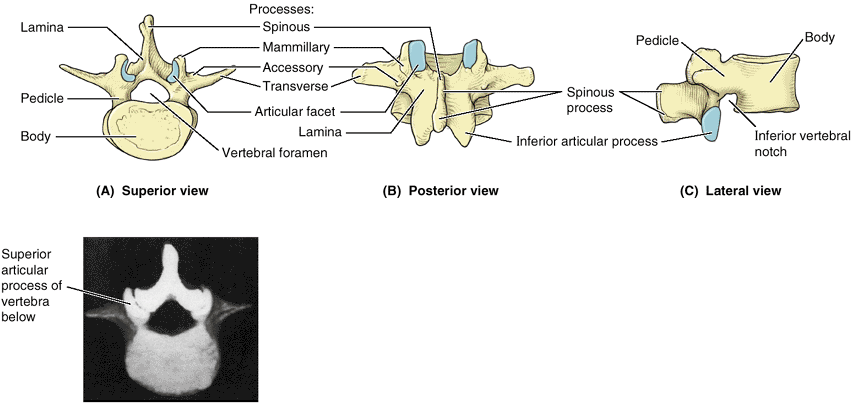
Figure 4.2. A “typical” vertebra, represented by L2. A. Functional components include the vertebral body (bone color), a vertebral arch (red), and seven processes: three for muscle attachment and leverage (blue) and four that participate in synovial joints with adjacent vertebrae (yellow). B and C.
Bony formations of the vertebrae are demonstrated. The vertebral foramen is bounded by the vertebral arch and body. A small superior vertebral notch and a larger inferior vertebral notch flank the pedicle. D. The superior and inferior notches of adjacent vertebrae plus the IV disc that unites them form the IV foramen for the passage of a spinal nerve and its accompanying vessels. Note that each articular process has an articular facet where contact occurs with the articular facets of adjacent vertebrae (B–D). |
characteristics from one region of the vertebral column to another and
to a lesser degree within each region; however, their basic structure
is the same. A typical vertebra (Fig. 4.2) consists of a vertebral body, a vertebral arch, and seven processes.1
more massive, roughly cylindrical, anterior part of the bone that gives
strength to the vertebral column and supports body weight. The size of
the vertebral bodies increases as the column descends, most markedly
from T4 inferiorly, as each bears progressively greater body weight.
(spongy, cancellous) bone enclosed by a thin external layer of compact
bone (Fig. 4.3). The trabecular bone is a
meshwork of mostly tall vertical trabeculae intersecting with short,
horizontal trabeculae. The interstices of these trabeculae are occupied
by red marrow that is among the most actively hematopoietic
(blood-forming) tissues of the mature individual. One or more large
foramina in the posterior surface of the body accommodate basivertebral
veins that drain the marrow (Fig. 4.20).
the vertebral body are covered with discs of hyaline cartilage
(vertebral “end plates”), which are remnants of the cartilaginous model
from which the bone develops (Bogduk, 1997). In dried
laboratory and museum skeletal specimens, this cartilage is absent, and
the exposed bone appears spongy, except at the periphery where an epiphysial rim or ring of smooth bone, derived from an anular epiphysis, is fused to the body (Fig. 4.2B).
In addition to serving as growth zones, the anular epiphyses and their
cartilaginous remnants provide some protection to the vertebral bodies
and permit some diffusion of fluid between the IV disc and the
capillaries in the vertebral body. The superior and inferior epiphyses
usually unite with the centrum, the primary ossification center for the central mass of the vertebral body (Fig. 4.2B), early in adult life (at approximately age 25) (see “Ossification of Vertebrae” in this chapter).
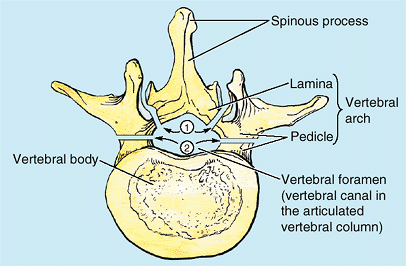
are short, stout cylindrical processes that project posteriorly from
the vertebral body to meet two broad, flat plates of bone, called laminae, which unite in the midline. The vertebral arch and the posterior surface of the vertebral body form the walls of the vertebral foramen (Fig. 4.2B & C). The succession of vertebral foramina in the articulated vertebral column forms the vertebral canal
(spinal canal), which contains the spinal cord and the roots of the
spinal nerves that emerge from it, along with the membranes (meninges),
fat, and vessels that surround and serve them (Fig. 4.1E). The vertebral notches
are indentations observed in lateral views of the vertebrae superior
and inferior to each pedicle between the superior and inferior
articular processes posteriorly and the corresponding projections of
the body anteriorly (Fig. 4.2C & D). The superior and inferior vertebral notches of adjacent vertebrae and the IV discs connecting them form the intervertebral foramina (Fig. 4.2D),
in which the spinal (posterior root) ganglia are located and through
which the spinal nerves emerge from the vertebral column with their
accompanying vessels.
-
One median spinous process
projects posteriorly (and usually inferiorly, typically overlapping the
vertebra below) from the vertebral arch at the junction of the laminae. -
Two transverse processes project posterolaterally from the junctions of the pedicles and laminae.
-
Four articular processes (G. zygapophyses)—two superior and two inferior—also arise from the junctions of the pedicles and laminae, each bearing an articular surface (facet).
transverse, afford attachments for deep back muscles and serve as
levers, facilitating the muscles that fix or change the position of the
vertebrae.
with corresponding processes of vertebrae adjacent (superior and
inferior) to them, forming zygapophysial (facet) joints (Fig. 4.2D).
Through their participation in these joints, these processes determine
the types of movements permitted and restricted between the adjacent
vertebrae of each region. The articular
processes
also assist in keeping adjacent vertebrae aligned, particularly
preventing one vertebra from slipping anteriorly on the vertebra below.
Generally, the articular processes bear weight only temporarily, as
when one rises from the flexed position, and unilaterally when the
cervical vertebrae are laterally flexed to their limit. However, the
inferior articular processes of the L5 vertebra bear weight even in the
erect posture.
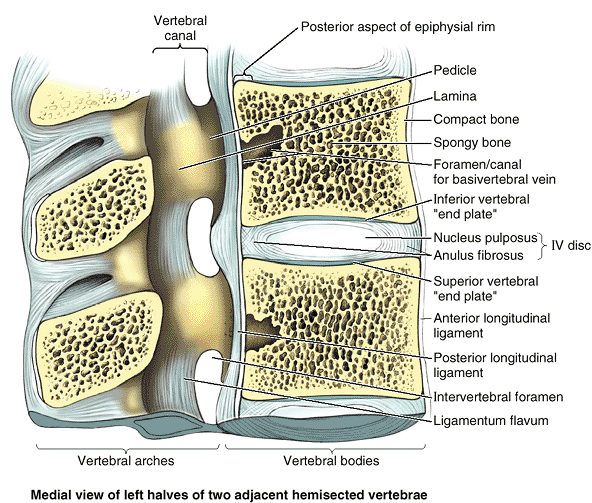 |
|
Figure 4.3. Internal aspects of vertebral body and vertebral canal.
Vertebral bodies consist largely of spongy bone, with tall, vertical supporting trabeculae linked by short horizontal trabeculae, covered by a relatively thin layer of compact bone. Hyaline cartilage end plates cover the superior and inferior surfaces of the bodies, surrounded by smooth bony epiphysial rims. The posterior longitudinal ligament, covering the posterior aspect of the vertebral bodies and linking the IV discs, forms the anterior wall of the vertebral canal. Lateral and posterior walls of the vertebral canal are formed by vertebral arches (pedicles and laminae) alternating with IV foramina and ligamenta flava. |
and increase in size proportionately, and vertebral arches, which
collectively house and protect the spinal cord. Processes extending
from the arch provide attachment and leverage for muscles or direct
movements between vertebrae.
and the adjacent supporting vertebral laminae in a particular region of
the vertebral column is called a laminectomy (1 in Fig. B4.1). The term is also commonly used to denote removal of most of the vertebral arch by transecting the pedicles (2 in Fig. B4.1).
Laminectomies are performed surgically (or anatomically in the
dissection laboratory) to gain access to the vertebral canal, providing
posterior exposure of the spinal cord (if performed above the L2 level)
and/or the roots of specific spinal nerves. Surgical laminectomy is
often performed to relieve pressure on the spinal cord or nerve roots
caused by a tumor, herniated IV disc, or bony hypertrophy (excess
growth).
 |
|
Figure B4.1
|
vertebrae demonstrate characteristic features identifying them as
belonging to one of the five regions of the vertebral column (e.g.,
vertebrae having foramina in their transverse processes are cervical
vertebrae). In addition, certain individual vertebrae have
distinguishing features; the C7 vertebra, for example, has the longest
spinous process. It forms a prominence under the skin at the back of
the neck, especially when the neck is flexed.
articular processes of the vertebrae in a characteristic direction that
determines the type of movement permitted between the adjacent
vertebrae and, in aggregate, for the region. For example, the articular
facets of thoracic vertebrae are nearly vertical, and together define
an arc centered in the IV disc; this arrangement permits rotation and
lateral flexion of the vertebral column in this region (Table 4.2). Regional variations in the size and shape of the vertebral canal accommodate the varying thickness of the spinal cord (Fig 4.1D & E).
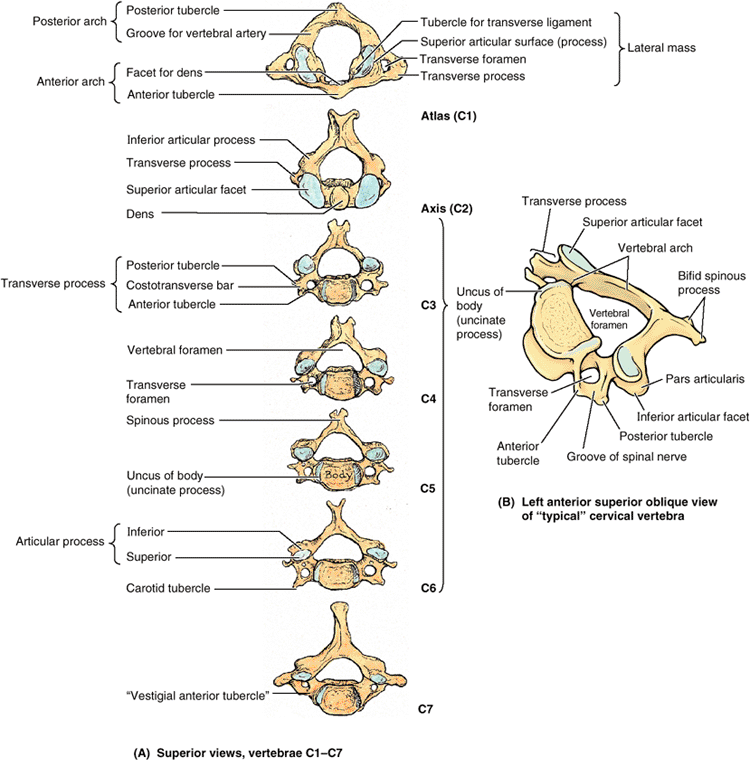
The smallest of the 24 movable vertebrae, the cervical vertebrae are
located between the cranium and the thoracic vertebrae. Their smaller
size reflects the fact that they bear less weight than do the larger
inferior vertebrae. Although the cervical IV discs are thinner than
those of inferior regions, they are relatively thick compared to the
size of the vertebral bodies they connect. The relative thickness of
the discs, the nearly horizontal orientation of the articular facets,
and the small amount of surrounding body mass give the cervical region
the greatest range and variety of movement of all the vertebral regions.
The vertebral arteries and their accompanying veins pass through the
transverse foramina, except those in C7, which transmit only small
accessory veins. Thus the foramina are smaller in C7 than those in
other cervical vertebrae, and occasionally they are absent. The
transverse processes of cervical vertebrae end laterally in two
projections: an anterior tubercle and a posterior tubercle. The tubercles provide attachment for a laterally placed group of cervical muscles (levator scapulae and scalenes). Grooves on the transverse processes between tubercles (the floor of the groove being formed by a costotransverse bar) accommodate the anterior rami of the cervical spinal nerves (Table 4.1B).
The carotid tubercles of vertebra C6 are so called carotid tubercles
because the common carotid arteries may be compressed here, in the
groove between the tubercle and body, to control bleeding from these
vessels. Bleeding may continue because of the carotid’s multiple
anastomoses of distal branches with adjacent and contralateral
branches, but at a slower rate.
|
Table 4.1. Cervical Vertebraea
|
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||||||
 |
|
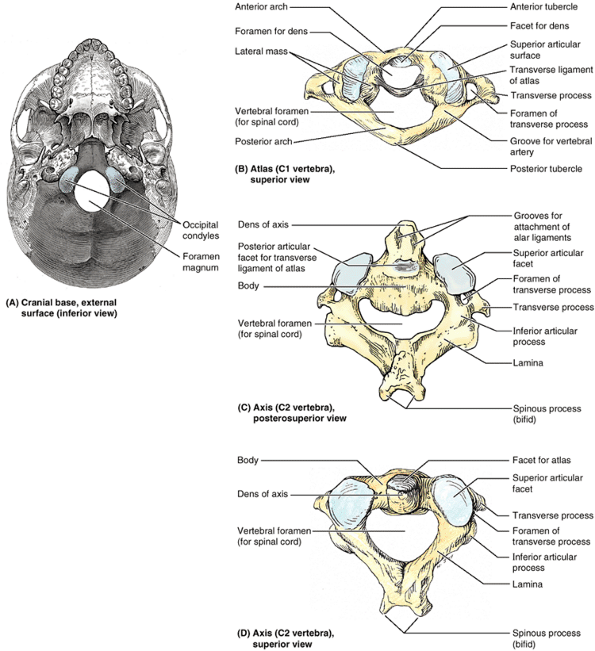
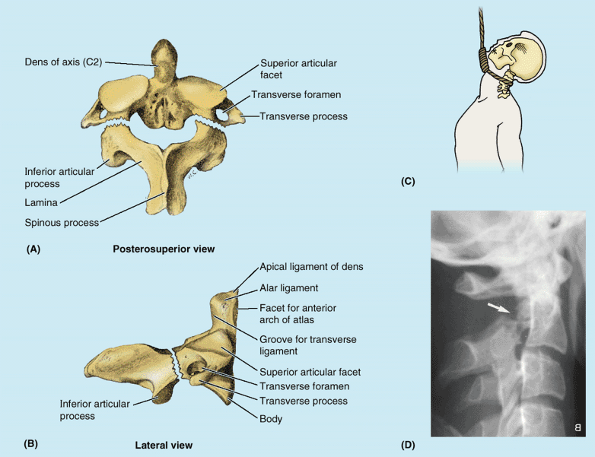
Figure 4.4. Cranial base and C1 and C2 vertebrae. A. Observe the occipital condyles that articulate with the superior articular surfaces (facets) of the atlas (vertebra C1). B.
The atlas, on which the cranium rests, has neither a spinous process nor a body. It consists of two lateral masses connected by anterior and posterior arches. C and D. The tooth-like dens characterizes the axis (vertebra C2) and provides a pivot around which the atlas turns and carries the cranium. It articulates anteriorly with the anterior arch of the atlas (“Facet for dens” in part B) and posteriorly with the transverse ligament of the atlas (see part B). |
They have large vertebral foramina to accommodate the enlargement of
the spinal cord in this region in relation to the innervation of the
upper limbs. The superior borders of the transversely elongated bodies
of the cervical vertebrae are elevated posteriorly and especially
laterally but are depressed anteriorly, resembling somewhat a sculpted
seat (Table 4.1B).
The inferior border of the body of the superiorly placed vertebra is
reciprocally shaped. The adjacent cervical vertebrae articulate in a
way that permits free flexion and extension and some lateral flexion
but restricted rotation. The planar, nearly horizontal articular facets
of the articular processes are also favorable for these movements. The
elevated superolateral margin is the uncus of the body (uncinate process).
The spinous processes of the C3–C6 vertebrae are short and usually
bifid in whites but usually not in people of African descent. C7 is a
prominent vertebra that is characterized by a long spinous process;
because of this prominent process, C7 is called the vertebra prominens. Run your finger along the midline of the posterior aspect of your neck until you feel the prominent C7 spinous process.
that serve the place of a body by bearing the weight of the globe-like
cranium in a manner similar to the way that Atlas of Greek mythology
bore the weight of the world on his shoulders. The transverse processes
of the atlas arise from the lateral masses, causing them to be more
laterally placed than those of the inferior vertebrae. This feature
makes the atlas the widest of the cervical vertebrae, thus providing
increased leverage for attached muscles. The kidney-shaped, concave superior articular surfaces of the lateral masses receive two large cranial protuberances called the occipital condyles at the sides of the foramen magnum (Fig. 4.4A). Anterior and posterior arches, each of which bears a tubercle in the center of its external aspect, extend between the lateral masses, forming a complete ring (Fig 4.4B). The posterior arch, which corresponds to the lamina of a typical vertebra, has a wide groove for the vertebral artery on its superior surface. The C1 nerve also runs in this groove.
is the strongest of the cervical vertebrae. C1, carrying the cranium,
rotates on C2, as when a person turns the head to indicate “no.” The
axis has two large, flat bearing surfaces, the superior articular facets, on which the atlas rotates (Fig. 4.4C). The distinguishing feature of the axis is the blunt tooth-like dens
(odontoid process), which projects superiorly from its body. Both the
dens (G. tooth) and the spinal cord inside its coverings are encircled
by the atlas. The dens lies anterior to the spinal cord and serves as
the pivot about which the rotation occurs. The dens is held in position
against the posterior aspect of the anterior arch of the atlas by the transverse ligament of the atlas (Fig. 4.4B).
This ligament extends from one lateral mass of the atlas to the other,
passing between the dens and spinal cord, forming the posterior wall of
the “socket” that receives the dens. Thus it prevents posterior
(horizontal) displacement of the dens and anterior displacement of the
atlas. Either displacement would compromise the portion of the
vertebral foramen of C1 that gives passage to the spinal cord. C2 has a
large bifid spinous process (Fig. 4.4C & D) that can be felt deep in the nuchal groove, the superficial vertical groove at the back of the neck.
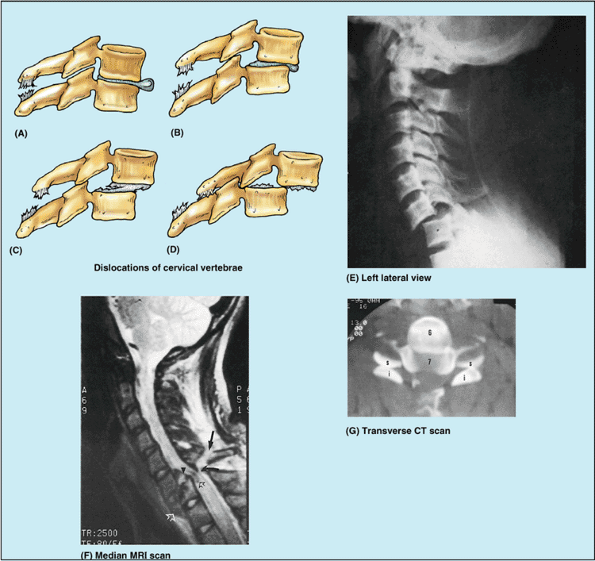
facets, the cervical vertebrae are less tightly interlocked than other
vertebrae. The cervical vertebrae, stacked like books, can be
dislocated in neck injuries with less force than is required to
fracture them (Fig. B4.2A–F).
Because of the large vertebral canal in the cervical region, slight
dislocation can occur here without damaging the spinal cord (Fig. B4.2B).
Severe dislocations, or dislocations combined with fractures
(fracture–dislocations) injure the spinal cord. If the dislocation does
not result in “facet jumping” with locking of the displaced articular
processes (Fig. B4.2F & G),
the cervical vertebrae may self-reduce (slip back into place) so that a
radiograph may not indicate that the cord has been injured. MRI,
however, may reveal the resulting soft tissue damage.
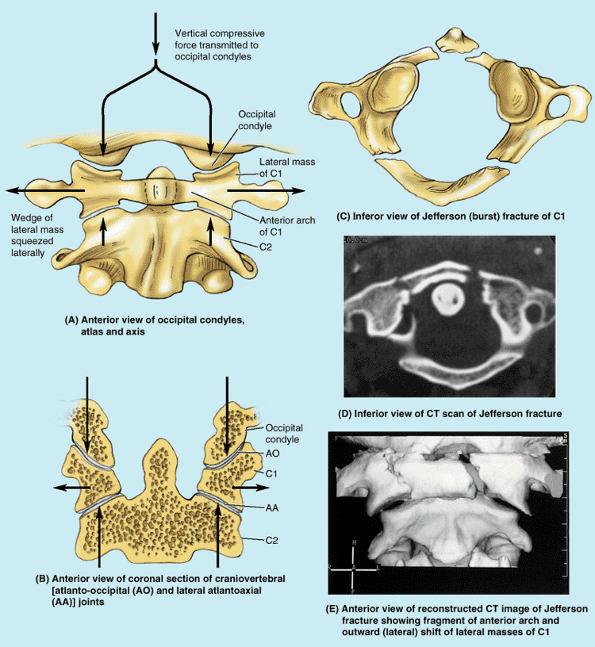
lateral masses connected by relatively thin anterior and posterior
arches and a transverse ligament (Fig. B4.3A).
Because the thick side of the lateral mass wedges is directed
laterally, vertical forces (as would result from a blow to the top of
the head from a falling object, or striking the bottom of a pool in a
diving accident) compressing the lateral masses between the occipital
condyles and the axis drive them apart, fracturing one or both of the
bony arches. If the force is sufficient, rupture of the transverse
ligament that links them will occur (Fig. B4.3B). The resulting Jefferson or burst fracture (Fig. B4.3C–E)
in itself does not necessarily result in spinal cord injury, because
the dimensions of the bony ring actually increase. Spinal cord injury
is more likely, however, if the transverse ligament has also been
ruptured (see clinical correlation [blue] box “Rupture of the Transverse Ligament of the Atlas,” in this chapter), indicated radiographically by widely spread lateral masses.
Usually the fracture occurs in the bony column formed by the superior
and inferior articular processes of the axis, the pars interarticularis
(Table 4.1B). A fracture in this location, called a traumatic spondylolysis of C2 (Fig. B4.4A, B, & D), usually occurs as a result of hyperextension of the head on the neck, rather than the combined hyperextension of the head and neck, which results in whiplash
injury. Such hyperextension of the head was used to execute criminals
by hanging, in which the knot was placed under the chin before the body
suddenly dropped its length through the gallows floor (Fig. B4.4C); thus this fracture is called a hangman’s fracture.
In more severe injuries, the body of the C2 vertebra is displaced
anteriorly with respect to the body of the C3 vertebra. With or without
such subluxation (incomplete dislocation) of the axis, injury of the
spinal cord and/or of the brainstem is likely, sometimes resulting in quadriplegia (paralysis of all four limbs) or death. Fractures of the dens are also common axis injuries (40–50%), which may result from a horizontal blow to the head or as a complication of osteopenia (pathological loss of bone mass) (see clinical correlation [blue] box “Fracture of the Dens,” in this chapter).
 |
|
Figure B4.2. Dislocations of cervical vertebrae. Four stages of injury are shown: (A) stage I, flexion sprain; (B) stage II, anterior subluxation with 25% anterior translation; (C) stage III, 50% translation; and (D) stage IV, complete dislocation. E. This lateral view radiograph shows a stage III dislocation with 50% translation. F. This MRI study of a stage IV dislocation with cord injury reveals that the body of C7 is fractured (open white arrowhead). The ligamentum flavum is disrupted (curved black arrow), and the spinous process is avulsed (straight black arrow). G. This transverse CT scan (same as shown in part F)
reveals the reversed position of the articular processes of the C6 and C7 vertebrae owing to “facet jumping.” The flat articular surfaces should be in contact, forming C6–C7 zygapophysial joints. |
 |
|
Figure B4.3
|
 |
|
Figure B4.4 Fracture and dislocation of axis. Posterosuperior (A) and lateral (B) views of a hangman’s fracture of the C2 vertebra are shown. C. This position of the knot produces hyperextension during hanging. D. This right lateral view radiograph reveals a hangman’s fracture (arrow) of C2.
|
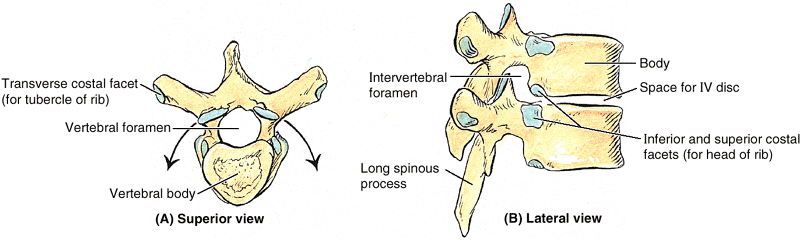
for articulation with ribs. The costal facets and other characteristic
features of thoracic vertebrae are illustrated and listed in Table 4.2.
The middle four thoracic vertebrae (T5–T8) demonstrate all the features
typical of thoracic vertebrae. The articular processes of thoracic
vertebrae extend vertically with paired, nearly coronally oriented
articular facets that define an arc centered in the IV disc. This arc
permits rotation and some lateral flexion of the vertebral column in
this region, in fact, the greatest degree of rotation is permitted here
(Table 4.2A).
Attachment of the rib cage combined with the vertical orientation of
articular facets and overlapping spinous processes limits flexion and
extension as well as lateral flexion.
vertebrae. T1 is atypical of thoracic vertebrae in that it has a long,
almost horizontal spinous process that may be nearly as prominent as
that of the vertebra prominens. T1 also has a complete costal facet on
the superior edge of its body for the 1st rib and a demifacet on its
inferior edge that contributes to the articular surface for the 2nd rib.
vertebrae, including tubercles similar to the accessory and mammillary
processes of lumbar vertebrae. However, most of the transition in
characteristics from thoracic to lumbar region occurs over the length
of a single vertebra: vertebra T12. Generally, its superior half is
thoracic in character, having costal facets and articular processes
that permit primarily rotatory movement, whereas its inferior half is
lumbar in character, devoid of costal facets and having articular
processes that permit only flexion and extension. Consequently,
vertebra T12 is subject to transitional stresses that cause it to be
the most commonly fractured vertebra.
|
Table 4.2. Thoracic Vertebrae
|
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||||
Because the weight they support increases toward the inferior end of
the vertebral column, lumbar vertebrae have massive bodies, accounting
for much of the thickness of the lower trunk in the median plane. Their
articular processes extend vertically, with articular facets sagittally
oriented initially (beginning abruptly with the T12–L1 joints) but
becoming more coronally oriented as the column descends. The L5–S1
facets are distinctly coronal in orientation. In the more sagittally
oriented superior joints, the laterally facing facets of the inferior
processes of the vertebra above are “gripped” by the medially facing
facets of the superior processes of the vertebra below, in a manner
that facilitates flexion and extension, allows lateral flexion, but
prohibits rotation (Figs. 4.1 and 4.2).
The transverse processes project somewhat posterosuperiorly as well as
laterally. On the posterior surface of the base of each transverse
process is a small accessory process,
which provides an attachment for the medial intertransverse lumborum
muscle. On the posterior surface of the superior articular processes
are mammillary processes, which give attachment to the multifidus and medial intertransverse muscles (back muscles).
carries the weight of the whole upper body. L5 is distinguished by its
massive body and transverse processes. Its body is markedly deeper
anteriorly; therefore, it is largely responsible for the lumbosacral
angle between the long axis of the lumbar region of the vertebral
column and that of the sacrum (Fig. 4.1D). Body weight is transmitted from L5 vertebra to the base of the sacrum, formed by the superior surface of S1 vertebra (Fig. 4.5A).
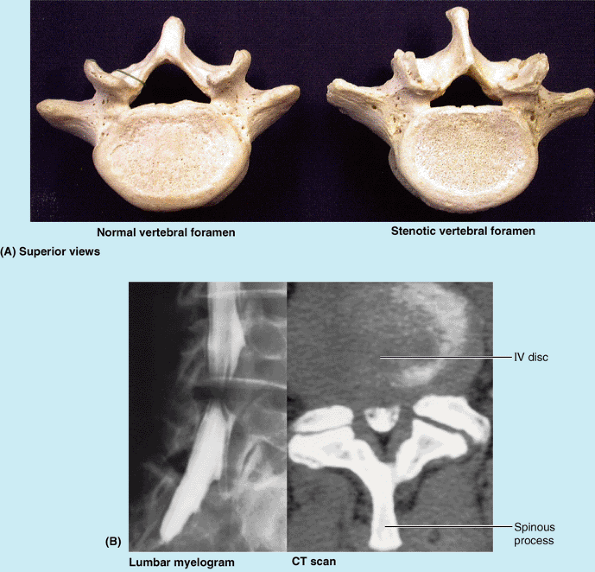
This condition may be a hereditary anomaly that can make a person more
vulnerable to age-related degenerative changes such as IV disc bulging
(Rowland and McCormick, 2000). The narrowing
is usually maximal at the level of the IV discs. However, stenosis of a
lumbar vertebral foramen alone may cause compression of one or more of
the spinal nerve roots occupying the inferior vertebral canal (Fig. 4.1E).
Electromyography can confirm that the denervation is restricted to
muscles innervated by the lumbosacral nerve roots. Surgical treatment
of lumbar stenosis may consist of decompressive laminectomy (see clinical correlation [blue] box “Laminectomy,” in this chapter). When IV disc protrusion occurs in a patient with spinal stenosis (Fig. B4.5B),
it further compromises a vertebral canal that is already limited, as
does arthritic proliferation and ligamentous degeneration (McCormick, 2000).
 |
|
Figure B4.5 Lumbar spinal stenosis. A. Normal and stenotic vertebral foramina are compared. B. The lumbar myelogram and CT scan demonstrate a high-grade stenosis caused by the IV disc bulging at the L4–L5 space.
|
It is located between the hip bones and forms the roof and
posterosuperior wall of the posterior pelvic cavity. The triangular
shape of the sacrum results from the rapid decrease in the size of the
lateral masses of the sacral vertebrae during development. The inferior
half of the sacrum is not weight bearing; therefore, its bulk is
diminished considerably. The sacrum (L. sacred or holy bone) provides
strength and stability to the pelvis and transmits the weight of the
body to the pelvic girdle, the bony ring formed by the hip bones and
sacrum, to which the lower limbs are attached.
(L. horse tail), that descend past the termination of the spinal cord.
On the pelvic and posterior surfaces of the sacrum between its
vertebral components are typically four pairs of sacral foramina for the exit of the posterior and anterior rami of the spinal nerves (Fig. 4.5A & B). The anterior (pelvic) sacral foramina are larger than the posterior (dorsal) ones.
formed by the superior surface of the S1 vertebra. Its superior
articular processes articulate with the inferior articular processes of
the L5 vertebra. The anterior projecting edge of the body of the S1
vertebra is the sacral promontory (L. mountain ridge), an important obstetrical landmark (see Chapter 3). The apex of the sacrum, its
tapering inferior end, has an oval facet for articulation with the coccyx.
|
Table 4.3. Lumbar Vertebrae
|
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||||
posterior part of the bony pelvis. The sacrum is tilted so that it
articulates with the L5 vertebra at the lumbosacral angle (Fig. 4.1D),
which varies from 130° to 160°. The sacrum is often wider in proportion
to length in the female than in the male, but the body of the S1
vertebra is usually larger in males.
Four transverse lines on this surface of sacra from adults indicate
where fusion of the sacral vertebrae occurred. During childhood, the
individual sacral vertebrae are connected by hyaline cartilage and
separated by IV discs. Fusion of the sacral vertebrae starts after age
20; however, most of the IV discs remain unossified up to or beyond
middle life (Williams et al., 1995).
represents the fused rudimentary spinous processes of the superior
three or four sacral vertebra; S5 has no spinous process. The intermediate sacral crests represent the fused articular processes, and the lateral sacral crests
are the tips of the transverse processes of the fused sacral vertebrae.
The clinically important features of the dorsal surface of the sacrum
are the inverted U-shaped sacral hiatus and the sacral cornua (L.
horns). The sacral hiatus results from the
absence of the laminae and spinous process of S5 and sometimes S4. The
sacral hiatus leads into the sacral canal. Its depth varies, depending
on how much of the spinous process and laminae of S4 are present. The sacral cornua,
representing the inferior articular processes of S5 vertebra, project
inferiorly on each side of the sacral hiatus and are a helpful guide to
its location.
It is the site of the synovial part of the sacroiliac joint between the
sacrum and ilium. During life, the auricular surface is covered with
hyaline cartilage.
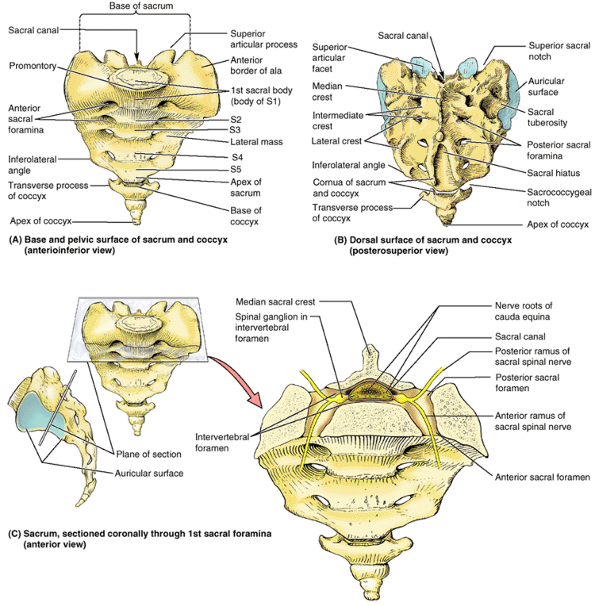 |
|
Figure 4.5. Sacrum and coccyx. A.
The base of the adult sacrum is the anterosuperior surface of the sacrum (i.e., the aspect of the sacrum opposite its apex). The base includes the articular surface of vertebra S1 (the anterior margin of which forms the sacral promontory), the sacral canal (the inferior part of the vertebral canal), and the right and left alae. Only the first of the four coccygeal vertebrae has transverse processes. B. The absence of the S4 and S5 spinous processes has resulted in the formation of a large sacral hiatus. The cornua, or horns, of the sacrum and coccyx are palpable clinical landmarks. C. Lateral and anterior orientation drawings of the sacrum in its anatomical position demonstrate the essentially frontal or coronal plane and level at which the sacrum has been sectioned to reveal the sacral canal containing the cauda equina. Spinal ganglia lie within the IV foramina, as they do at superior vertebral levels. However, the sacral posterior and anterior rami of the spinal nerves exit via posterior and anterior (pelvic) sacral foramina, respectively. The lateral orientation drawing demonstrates the auricular surface that joins the ilium to form the synovial part of the sacroiliac joint. In the anatomical position, the S1–S3 vertebrae lie in an essentially transverse plane, forming a roof for the pelvic cavity. |
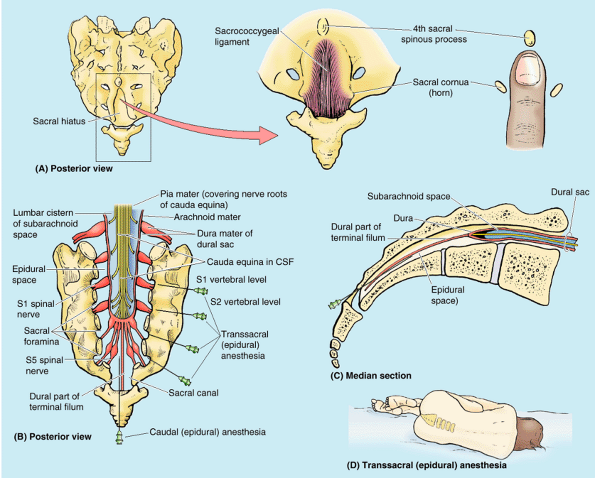
(a connective tissue strand extending from the tip of the spinal cord
to the coccyx). Deep (superior) to the ligament, the epidural space of
the sacral canal is filled with fatty connective tissue (Fig. B4.6A). In caudal epidural anesthesia or caudal analgesia,
a local anesthetic agent is injected into the fat of the sacral canal
that surrounds the proximal portions of the sacral nerves. This can be
accomplished by several routes, including the sacral hiatus (Fig. B4.6B & C).
Because the sacral hiatus is located between the sacral cornua and
inferior to the S4 spinous process or median sacral crest, these
palpable bony landmarks are important for locating the hiatus (Fig. B4.6A).
The anesthetic solution spreads superiorly and extradurally, where it
acts on the S2–Co spinal nerves of the cauda equina. The height to
which the anesthetic ascends is controlled by the amount injected and
the position of the patient. Sensation is lost inferior to the epidural
block. Anesthetic agents can also be injected through the posterior
sacral foramina into the sacral canal around the spinal nerve roots (transsacral epidural anesthesia) (Fig. B4.6B). The role of epidural anesthesia during childbirth and the areas anesthetized are discussed in Chapter 3.
 |
|
Figure B4.6
|
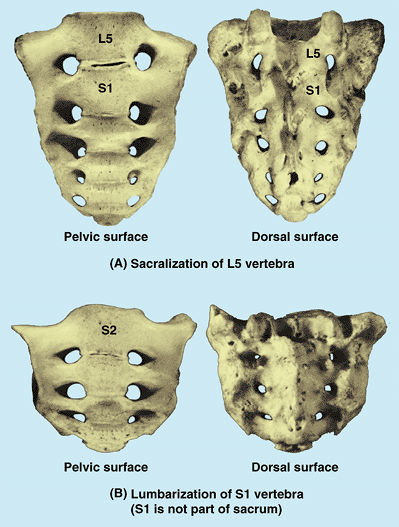
respectively. In others, S1 is more or less separated from the sacrum
and is partly or completely fused with L5 vertebra, which is called lumbarization of the S1 vertebra (Fig. B4.7B). When L5 is sacralized, the L5–S1 level is strong and the L4–L5 level degenerates, often producing painful symptoms.
 |
|
Figure B4.7
|
small triangular bone that is usually formed by fusion of the four
rudimentary coccygeal vertebrae, although in some people, there may be
one less or one more (Fig. 4.5). The coccygeal
vertebra 1 (Co1) may be separate. The coccyx is the remnant of the
skeleton of the embryonic tail-like caudal eminence, which is present
in human embryos from the end of the 4th week until the beginning of
the 8th week (Moore and Persaud, 2003). The
pelvic surface of the coccyx is concave and relatively smooth, and the
posterior surface has rudimentary articular processes. Co1 is the
largest and broadest of all the coccygeal vertebrae. Its short
transverse processes are connected to the sacrum, and its rudimentary
articular processes form coccygeal cornua,
which articulate with the sacral cornua. The last three coccygeal
vertebrae often fuse during middle life, forming a beak-like coccyx;
this accounts for its name (G. coccyx,
cuckoo). With increasing age, Co1 often fuses with the sacrum, and the
remaining coccygeal vertebrae usually fuse to form a single bone. The
coccyx does not participate with the other vertebrae in support of the
body weight when standing; however, when sitting it may flex anteriorly
somewhat, indicating that it is receiving some weight. The coccyx
provides attachments for parts of the gluteus maximus and coccygeus
muscles and the anococcygeal ligament, the median fibrous intersection of the pubococcygeus muscles (see Chapter 3).
subperiosteal bruising or fracture of the coccyx, or a
fracture–dislocation of the sacrococcygeal joint. Displacement is
common, and surgical removal of the fractured bone may be required to
relieve pain. An especially difficult childbirth occasionally injures
the mother’s coccyx. A troublesome syndrome, coccygodynia, often follows coccygeal trauma; pain relief is commonly difficult.
transverse foramina for cervical vertebrae, (2) costal facets for
thoracic vertebrae, (3) the absence of transverse foramina and costal
facets for lumbar vertebrae, (4) the fusion of adjacent sacral
vertebrae, and (5) the rudimentary nature of coccygeal vertebrae.
Later, these mesenchymal bone models chondrify and cartilaginous
vertebrae form. Typically, vertebrae begin to ossify toward the end of
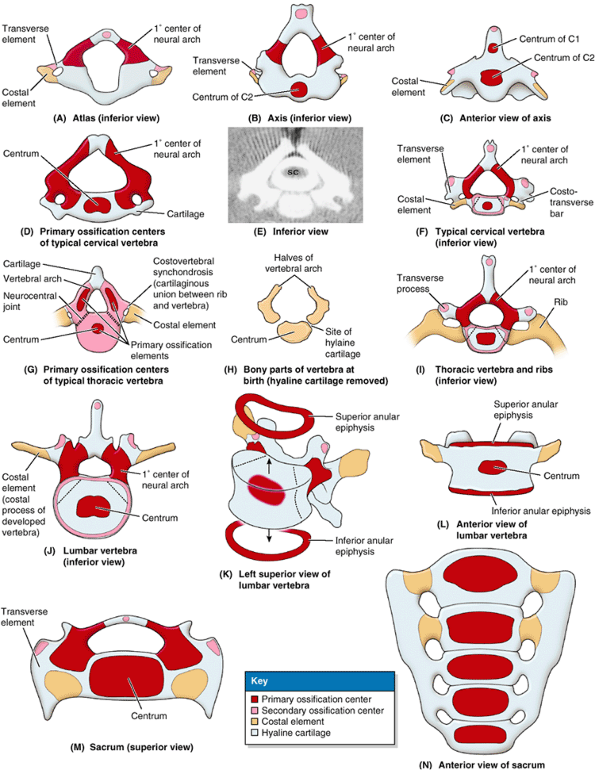
the embryonic period (8th week), with three primary ossification centers develop
ing in each cartilaginous vertebra: an endochondral centrum, which will eventually constitute most of the body of the vertebra, and two perichondral centers, one in each half of the neural arch (Fig. 4.6A–J & M).
Ossification continues throughout the fetal period. At birth, each
typical vertebra and the superiormost sacral vertebrae consists of
three bony parts united by hyaline cartilage. The inferior sacral
vertebrae and all the coccygeal vertebrae are still entirely
cartilaginous; they ossify during infancy. The halves of the neural
arches articulate at neurocentral joints, which are primary cartilaginous joints (Fig. 4.6G).
The halves of the neural/vertebral arch begin to fuse with each other
posterior to the vertebral canal during the 1st year, beginning in the
lumbar region and then in the thoracic and cervical regions. The neural
arches begin fusing with the centra in the upper cervical region around
the end of the 3rd year, but usually the process is not completed in
the lower lumbar region until after the 6th year.
develop during puberty in each typical vertebra: one at the tip of the
spinous process; one at the tip of each transverse process; and two
anular epiphyses (ring epiphyses), one on the superior and one on the
inferior edges of each vertebral body (i.e., around the margins of the
superior and inferior surfaces of the vertebral body) (Fig. 4.6F & I–L). The hyaline anular epiphyses, to which the IV discs attach, are sometimes referred to as epiphysial growth plates and form the zone from which the vertebral body grows in height (see “Cartilage and Bones” in the Introduction).
When growth ceases early in the adult period, the epiphyses usually
unite with the vertebral body. This union results in the characteristic
smooth raised margin, the epiphysial rim, around the edges of the superior and inferior surfaces of the body of the adult vertebra (Figs. 4.2B and 4.3).
All secondary ossification centers have usually united with the
vertebrae by age 25; however, the ages at which specific unions are
made vary.
The costal elements normally develop into ribs only in the thoracic
region; they become part of the transverse process or its equivalent at
other levels.
Also as a result of the cervical transverse processes being formed from
the two developmental elements, the transverse processes of cervical
vertebrae end laterally in an anterior tubercle (from the costal element) and a posterior tubercle
(from the transverse element). The atypical morphology of vertebrae C1
and C2 is also established during development. The centrum of C1
becomes fused to that of C2 and loses its peripheral connection to the
remainder of C1, thus forming the dens (Fig. 4.6C).
Since these first two centra are fused and are now part of C2, no IV
disc is formed between C1 and C2 to connect them together. The part of
the body that remains with C1 is represented by the anterior arch and tubercle of C1.
from the developing vertebrae and elongate into ribs, and the
transverse elements alone form the transverse processes (Fig. 4.6I).
by the fusion of the transverse and costal elements. (For more
information about the ossification of vertebrae, see Williams et al., 1995.)
In 1–2% of people, the developmental costal element of C7, which
normally becomes a small part of the transverse process that lies
anterior to the transverse foramen (Fig. 4.6F),
becomes abnormally enlarged. This structure may vary in size from a
small protuberance to a complete rib that occurs bilaterally about 60%
of the time. The supernumerary (extra) rib or a fibrous connection
extending from its tip to the first thoracic rib may elevate and place
pressure on structures that emerge from the superior thoracic aperture:
notably the subclavian artery or inferior trunk of the brachial plexus
and may cause thoracic outlet syndrome (see clinical correlation [blue] box “Supernumerary Ribs,” in Chapter 1).
vertebra increases in height threefold (from 5–6 mm to 15–18 mm), and
between ages 5 and 13, it increases another 45–50%. Longitudinal growth
continues throughout adolescence, but the rate decreases and is
completed between ages 18 and 25. During middle and older age, there is
an overall decrease in bone density and strength, particularly
centrally within the vertebral body. Consequently, the articular
surfaces gradually bow inward, so that both the superior and inferior
surfaces of the vertebrae become increasingly concave (Fig. B4.8A)
and the IV discs become increasingly convex. The bone loss and
consequent change in shape of the vertebral bodies may account in part
for the slight loss in height that occurs with aging.
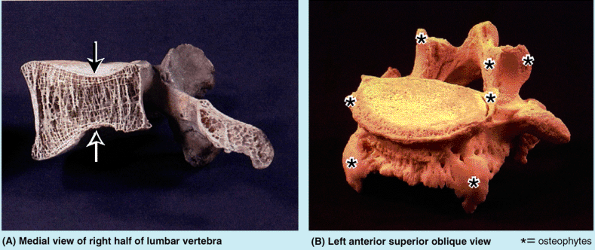
apparent narrowing of the intervertebral “space” on radiographs based
on the distance between the margins of the vertebral bodies; however,
this should not be interpreted as a loss of IV disc thickness. Aging of
the IV discs combined with the changing shape of the vertebrae results
in an increase in compressive forces at the periphery of the vertebral
bodies, where the discs attach. In response, osteophytes (bony spurs)
commonly develop around the margins of the vertebral body (along the
attachments of the fibers outer part of the disc), especially
anteriorly and posteriorly (Fig. B4.8B).
Similarly, as altered mechanics place greater stresses on the
zygapophysial joints, osteophytes develop along the attachments of the
joint capsules and accessory ligaments, especially those of the
superior articular process, whereas extensions of the articular
cartilage (“wraparound bumpers”) develop around the articular facets of
the inferior processes. This bony or cartilaginous growth during
advanced age has traditionally been viewed as a disease process (spondylosis in the case of the vertebral bodies, osteoarthrosis
in the case of the zygapophysial joints), but it may be more realistic
to view it as an expected morphological change with age, representing normal anatomy for a particular age range.
difficult: Some patients with these manifestations present with pain,
others demonstrate the same age-related changes but have no pain, and
still others exhibit little morphological change but complain of the
same types of pain as those with evident change. In view of this and
the typical occurrence of these findings, some have suggested that
consideration of such age-related changes as pathological processes is
not justified (Bogduk, 1997).
fuse. Caution must be exercised so that a persistent epiphysis is not
mistaken for a vertebral fracture in a radiograph or CT scan.
in which the laminae (neural arches) of L5 and/or S1 fail to develop
normally and fuse posterior to the vertebral canal. This bony defect,
present in up to 24% of the population (Greer, 2000),
usually occurs in the vertebral arch of L5 and/or S1. The defect is
concealed by the overlying skin, but its location is often indicated by
a tuft of hair (Moore and Persaud, 2003). Most
people with spina bifida occulta have no back problems. When examining
a newborn, adjacent vertebrae should be palpated in sequence to be
certain the vertebral arches are intact and continuous from the
cervical to the sacral regions.
one or more vertebral arches may fail to develop completely. Spina
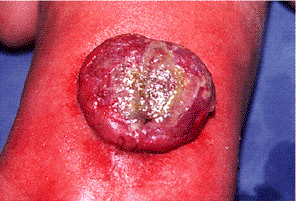
bifida cystica is associated with herniation of the meninges (meningocele, a spina bifida associated with a meningeal cyst) and/or the spinal cord (meningomyelocele) (Fig. B4.9).
Neurological symptoms are usually present in severe cases of
meningomyelocele (e.g., paralysis of the limbs and disturbances in
bladder and bowel control). Severe forms of spina bifida result from neural tube defects, such as the defective closure of the neural tube during the 4th week of embryonic development (Moore and Persaud, 2003).
 |
|
Figure B4.8. Effects of aging on vertebrae.
|
 |
|
Figure B4.9. Posterior view of spina bifida cystica.
|
may result in 32 or 34 vertebrae. Estimates of the frequency of
abnormal numbers of vertebrae superior to the sacrum (the normal number
is 24) range between 5% and 12%. Variations in vertebrae are affected
by race, sex, and developmental factors (genetic and environmental). An
increased number of vertebrae occurs more often in males and a reduced
number occurs more frequently in females. Some races show more
variation in the number of vertebrae. Variations in the number of
vertebrae may be clinically important: An increased length of the
presacral region of the vertebral column increases the strain on the
inferior part of the lumbar region of the column owing to the increased
leverage. However, most numerical variations are detected incidentally
during diagnostic medical imaging studies being performed for other
reasons and during dissections and autopsies of persons with no history
of back problems.
(e.g., when reporting the site of a vertebral fracture). When counting
the vertebrae, begin at the base of the neck. The number of cervical vertebrae (seven) is remarkably constant
(and not just in humans, but among mammals—even giraffes and snakes
have seven cervical vertebrae!). When considering a numerical
variation, the thoracic and lumbar regions must be considered together
because some people have more than five lumbar vertebrae and a
compensatory decrease in the number of thoracic vertebrae (O’Rahilly, 1986).
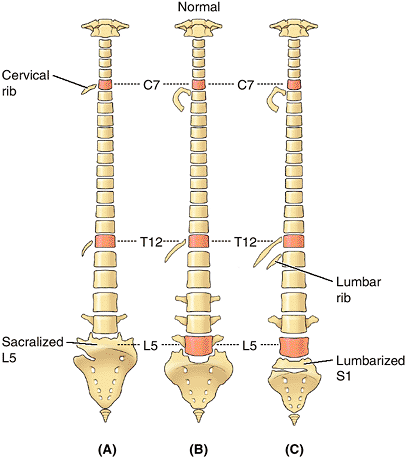
between the vertebrae and ribs, and the number of vertebrae that fuse
to form the sacrum (Fig. 4.7). The relationship of presacral vertebrae to ribs and/or sacrum may occur higher (cranial shift) or lower (caudal shift)
than normal. Note, however, that a C7 vertebra articulating with a
rudimentary cervical rib(s) is still considered a cervical vertebra.
The same is true for lumbar vertebrae and lumbar ribs. Likewise, an L5
vertebra fused to the sacrum is referred to as a “sacralized 5th lumbar
vertebrae” (see clinical correlation [blue] box “Abnormal Fusion of Vertebrae,” in this chapter).
ossification centers within a cartilaginous model: a centrum that will
form most of the body and a center in each half of the neural arch.
Thus by the time of birth, most vertebrae consist of three bony parts
united by hyaline cartilage. Fusion occurs during the first 6 years in
a centrifugal pattern from the lumbar region. During puberty, five
secondary ossification centers appear: three related to the spinous and
transverse processes, and two anular epiphyses around the superior and
inferior margins of the vertebral body. Costal elements formed in
association with the ossification center of the transverse process
usually form ribs only in the thoracic region. They form components of
the transverse processes or their equivalents in other regions.
Knowledge of the pattern of ossification of the vertebrae allows
understanding of the normal structure of typical and atypical
vertebrae, as well as variations and malformations.
aggregate structure, normally made up of all 33 vertebrae and the
components that unite them into a single structural, functional
entity—the “axis” of the axial skeleton. Because it provides the
semirigid, central “core” about which movements of the trunk occur,
“soft” or hollow structures that run a longitudinal course are subject
to damage or kinking (e.g., the spinal cord, descending aorta, venae
cavae, thoracic duct, and esophagus). Thus they lie in close proximity
to the vertebral axis, where they receive its semirigid support and
torsional stresses on them are minimized.
-
Joints of the vertebral bodies.
-
Joints of the vertebral arches.
-
Craniovertebral (atlantoaxial and atlanto-occipital) joints.
-
Costovertebral joints (see Chapter 1).
-
Sacroiliac joints (see Chapter 3).
 |
|
Figure 4.6. Ossification of vertebrae.
Typical vertebrae develop from three primary ossification centers that are present at birth and five secondary ossifications centers that appear at puberty. Costal elements are usually incorporated into the transverse processes, except in the thoracic region where they become ribs. A. Vertebra C1 (atlas) lacks a centrum. B and C. Vertebra C2 (axis) has two centra, one of which forms most of the dens. D–F. The development of “typical” cervical vertebrae is shown, including (D) the primary ossification centers within the hyaline cartilage, (E) a CT scan of the vertebra shown in part D (SC, spinal cord), and (F) the primary and secondary ossification centers. G–I. The development of thoracic vertebrae is shown, including (G) the three primary ossification centers in a cartilaginous vertebra of a 7-week-old embryo (observe the joints present at this stage), (H) the bony parts of a thoracic vertebra after skeletonization (cartilage removed), and (I) the primary and secondary ossification centers (with ribs developed from costal elements). J–L. The development of the lumbar vertebrae is shown, including (J) the primary and secondary ossification centers, (K) the anular epiphyses separated from the body, and (L) the anular epiphyses in place. M and N. The development of the sacrum is shown. Note that the ossification and fusion of sacral vertebrae may not be completed until age 35. |
 |
|
Figure 4.7. Variations in vertebrae and their relationship to ribs. A.
A “cranial shift” is demonstrated, in which there are 13 ribs, including a cervical rib articulating with vertebra C7 and a diminished 12th rib articulating with vertebra T12. Vertebra L5 is shown partially incorporated into the sacrum, but such “sacralization” can also be complete. The lowest sacral segment (S5) is partially segmented. B. The common arrangement of the vertebrae and the position of 1st and 12th ribs are shown. C. A “caudal shift” is shown, in which the 12th rib is increased in size, and there is a small lumbar rib. The transverse process of vertebra L4 is increased in size, whereas those of vertebra L5 are greatly reduced. The first sacral segment is shown partially separated from the rest of the sacrum, but such “lumbarization” can also be complete. The 1st coccygeal segment is incorporated into the sacrum—that is, it is “sacralized.” |
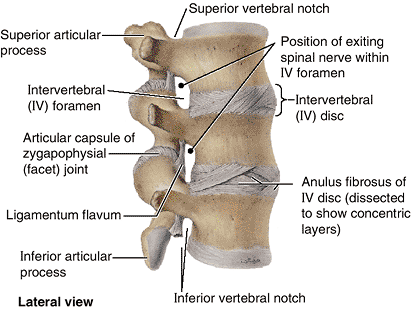
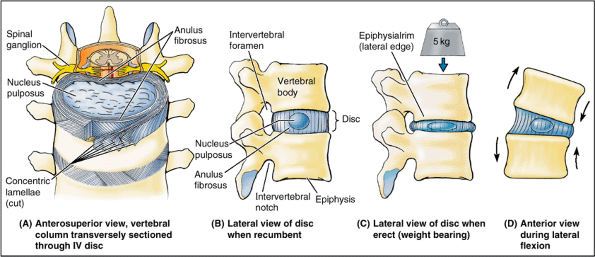
The IV discs provide strong attachments between the vertebral bodies,
uniting them into a continuous semirigid column and forming the
inferior half of the anterior border of the IV foramen. In aggregate,
the discs account for 20–25% of the length (height) of the vertebral
column (Fig. 4.1). As well as permitting
movement between adjacent vertebrae, their resilient deformability
allows them to serve as shock absorbers. Each IV disc consists of an anulus fibrosus, an outer fibrous part, composed of
concentric lamellae of fibrocartilage, and a gelatinous central mass, called the nucleus pulposus.
 |
|
Figure 4.8 Lumbar vertebrae and IV discs.
This view of the superior lumbar region shows the structure of the anuli fibrosi of the discs and the structures involved in formation of IV foramina. The disc forms the inferior half of the anterior boundary of an IV foramen (except in the cervical region). Thus herniation of the disc will not affect the spinal nerve exiting from the superior bony part of that foramen. |
The fibers forming each lamella run obliquely from one vertebra to
another; the fibers of one lamella typically run at right angles to
those of the adjacent ones. This arrangement, allows some movement
between adjacent vertebrae, while providing a strong bond between them.
At birth, these pulpy nuclei are about 88% water and are initially more
cartilaginous than fibrous. Their semifluid nature is responsible for
much of the flexibility and resilience of the IV disc and of the
vertebral column as a whole. Vertical forces deform the discs, which
thus serve as shock absorbers. The pulpy nuclei become broader when
compressed and thinner when tensed or stretched (as when hanging or
suspended) (Fig. 4.9C).
Compression and tension occur simultaneously in the same disc during
anterior and lateral flexion and extension of the vertebral column (Fig. 4.9D).
During these movements, as well as during rotation, the turgid nucleus
acts as a semifluid fulcrum. Because the lamellae of the anulus
fibrosus are thinner and less numerous posteriorly than they are
anteriorly or laterally, the nucleus pulposus is not centered in the
disc but is more posteriorly placed. The nucleus pulposus is avascular;
it receives its nourishment by diffusion from blood vessels at the
periphery of the anulus fibrosus and vertebral body.
most inferior functional disc is between L5 and S1 vertebrae. The discs
vary in thickness in different regions; they are thickest relative to
the size of the bodies they connect in the cervical and lumbar regions
(and absolutely thickest in the latter) and thinnest in the superior
thoracic region. Their relative thickness is clearly related to the
range of movement, and their varying shapes produce the secondary
curvatures of the vertebral column. The discs are thicker anteriorly in
the cervical and lumbar regions, and their thickness is most uniform in
the thoracic region.
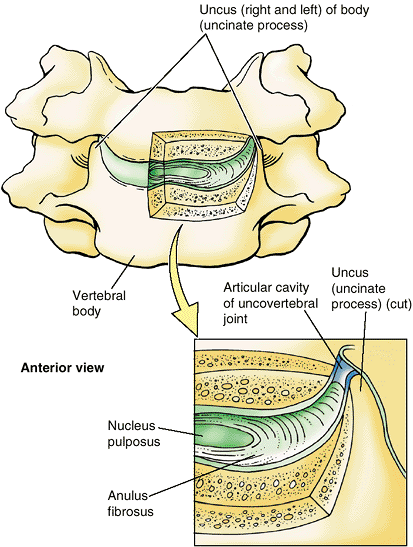
Luschka) are located between the unci of the bodies of C3–C6 vertebrae
and the beveled inferolateral surfaces of the vertebral bodies superior
to them (Fig. 4.10). The joints are at the
lateral and posterolateral margins of the IV discs. The articulating
surfaces of these joint-like structures are covered with cartilage
moistened by fluid contained within an interposed potential space, or
“capsule.” They are considered synovial joints by some; others consider
them to be degenerative spaces (fissures) in the discs occupied
by extracellular fluid. The uncovertebral joints are frequent sites of spur formation, which may cause neck pain.
 |
|
Figure 4.9. Structure and function of IV discs. A.
The disc consists of a nucleus pulposus and an anulus fibrosus. The superficial layers of the anulus have been cut and spread apart to show the direction of the fibers. Note that the combined thickness of the rings of the anulus is diminished posteriorly—that is, the anulus is thinner posteriorly. B. The fibrogelatinous nucleus pulposus occupies the center of the disc and acts as a cushion and shock-absorbing mechanism. C. The pulpy nucleus flattens and the anulus bulges when weight is applied, as occurs during standing and more so during lifting. D. During flexion and extension movements, the nucleus pulposus serves as a fulcrum. The anulus is simultaneously placed under compression on one side and tension on the other. |
 |
|
Figure 4.10. Uncovertebral joints.
These small, synovial joint-like structures are between the unci of the bodies of the lower vertebrae and the beveled surfaces of the vertebral bodies superior to them. These joints are at the posterolateral margins of the IV discs. |
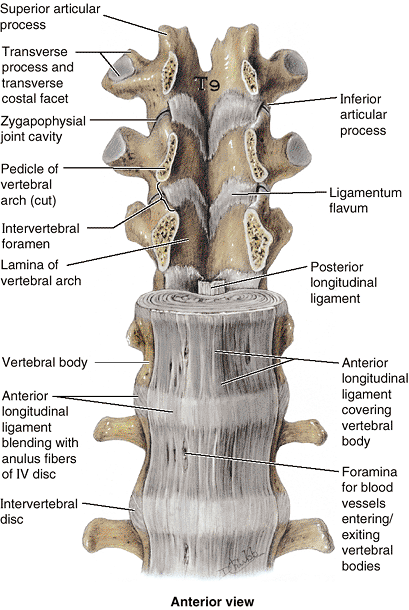
The ligament extends from the pelvic surface of the sacrum to the
anterior tubercle of vertebra C1 and the occipital bone anterior to the
foramen magnum. This ligament prevents hyperextension of the vertebral
column, maintaining stability of the joints between the vertebral
bodies. The anterior longitudinal ligament is the only ligament that
limits extension; all other IV ligaments limit forms of flexion.
The posterior longitudinal ligament runs within the vertebral canal
along the posterior aspect of the vertebral bodies. It is attached
mainly to the IV discs and less so to the posterior aspects of the
vertebral bodies from C2 to the sacrum, often bridging fat and vessels
between the ligament and the bony surface. This ligament weakly resists
hyperflexion of the vertebral column and helps prevent or redirect
posterior herniation of the nucleus pulposus. It is well provided with
nociceptive (pain) nerve endings.
 |
|
Figure 4.11. Relationship of ligaments to vertebrae and IV discs.
The lower thoracic (T9–T12) and upper lumbar (L1–L2) vertebrae, with associated discs and ligaments, are shown. The pedicles of the T9–T11 vertebrae have been sawn through and their bodies and intervening discs removed to provide an anterior view of the posterior wall of the vertebral canal formed by the laminae of the vertebral arches and the ligamenta flava extending between them. Between the adjacent left or right pedicles, the inferior and superior articular processes and the zygapophysial joints between them (from which joint capsules have been removed) and the lateralmost extent of the ligamenta flava form the posterior boundaries of IV foramina. The anterior longitudinal ligament is broad, whereas the posterior longitudinal ligament is narrow. Although thickest on the anterior aspect of the vertebral bodies (and often only this thickest part is illustrated), the anterior longitudinal ligament covers both the anterior and the lateral aspects of the bodies to the IV foramen. |
and proteoglycans while gaining collagen. As a result, the IV discs
lose their turgor, becoming stiffer and more resistant to deformation.
As the nucleus dehydrates and gains collagen, the two parts of the disc
appear to merge as the distinction between them becomes increasingly
diminished. With advancing age, the nucleus becomes dry and granular
and may disappear altogether as a distinct formation. As this occurs,
the anulus fibrosis assumes an increasingly greater share of the
vertical load and the stresses and strains that come with it. The
lamellae of the anulus thicken with age and often develop fissures and
cavities.
usually described as typical of the aging process and said to account
for loss of trunk stature. Although the margins of adjacent vertebral
bodies may approach more closely as the superior and inferior surfaces
of the body become shallow concavities (the most probable reason for
slight loss of height with aging), it has been shown that the
intervertebral bodies increase in size
with age. Not only do they become increasing convex but, between the
ages of 20 and 70, their anteroposterior (AP) diameter increases about
10% in females and 2% in males, while thickness (height) increases
centrally about 10% in both sexes. Thus disc height (as measured in the
center of the disc) is normally maintained with aging; overt or marked
cases of disc narrowing, especially when it is greater than that of
more superiorly located discs, suggests pathology, not normal aging (Bogduk, 1997).
pulposus into or through the anulus fibrosus is a well-recognized cause
of low back and lower limb pain (Fig. B4.10A & D).
The IV discs in young persons are strong—usually so strong that the
vertebrae often fracture during a fall before the discs rupture.
Furthermore, the water content of their nuclei pulposi is high
(approaching 90%), giving them great turgor (fullness). However,
violent hyperflexion of the vertebral column may rupture an IV disc and
fracture the adjacent vertebral bodies. Flexion of the vertebral column
produces compression anteriorly and stretching or tension posteriorly,
squeezing the nucleus pulposus further posteriorly toward the thinnest
part of the anulus fibrosus. If the anulus fibrosus has degenerated,
the nucleus pulposus may herniate into the vertebral canal and compress
the spinal cord (Fig. B4.10B & C) or the nerve roots of the cauda equina (Fig. B4.10A & D).
A herniated IV disc is inappropriately called a “slipped disc” by some
people. Sports writers often call the injury a “ruptured disc.”
posterolaterally, where the anulus fibrosus is relatively thin and does
not receive support from either the posterior or the anterior
longitudinal ligaments. A posterolateral herniated IV disc is more
likely to be symptomatic because of the proximity of the spinal nerve
roots. The localized back pain of a
herniated disc, which is usually acute, results from pressure on the
longitudinal ligaments and periphery of the anulus fibrosus and from
local inflammation caused by chemical irritation by substances from the
ruptured nucleus pulposus. Chronic pain
resulting from compression of the spinal nerve roots by the herniated
disc is referred to the area (dermatome) supplied by that nerve.
Because the IV discs are largest in the lumbar and lumbosacral regions,
where movements are consequently greater, posterolateral herniations of
the nucleus pulposus are most common here. Approximately 95% of lumbar
disc protrusions occur at the L4–L5 or L5–S1 levels. The marked
decrease in the radiographic intervertebral space (i.e., in disc
height) that may occur as a result of acute herniation of a nucleus may
also result in narrowing of the IV foramina, perhaps exacerbating the
compression of the spinal nerve roots, especially if hypertrophy of the
surrounding bone has also occurred. Because the nucleus pulposus
becomes increasingly dehydrated and fibrous or even granular or solid
with aging, a diagnosis of acute herniation in a patient of advanced
years should be regarded with suspicion. Rather, it is likely that the
nerve roots are being compressed by increased ossification of the IV
foramen as they exit.
low back pain, may be caused by a mild posterolateral protrusion of a
lumbar IV disc at the L5–S1 level that affects nociceptive (pain)
endings in the region, such as those associated with the posterior
longitudinal ligament. The clinical picture varies considerably, but
pain of acute onset in the lower back is a common presenting symptom.
Because muscle spasm is associated with low back pain, the lumbar
region of the vertebral column becomes tense and increasingly cramped
as a relative ischemia occurs, causing painful movement. With
treatment, the lumbago type of back pain usually begins to fade after a
few days; however, it may gradually be replaced by sciatica.
back and hip radiating down the back of the thigh into the leg, is
often caused by a herniated lumbar IV disc that compresses and
compromises the L5 or S1 component of the sciatic nerve. The IV
foramina in the lumbar region decrease in size and the lumbar nerves
increase in size, which may explain why sciatica is so common. New bone
(osteophytes) deposited around the zygapophysial joints or the
posterolateral margins during aging may narrow the foramina even more,
causing shooting pains down the lower limbs. Any maneuver that
stretches the sciatic nerve, such as flexing the thigh with the knee
extended (Kernig maneuver), may produce or exacerbate (but in some
individuals relieves) the pain caused by disc herniation.
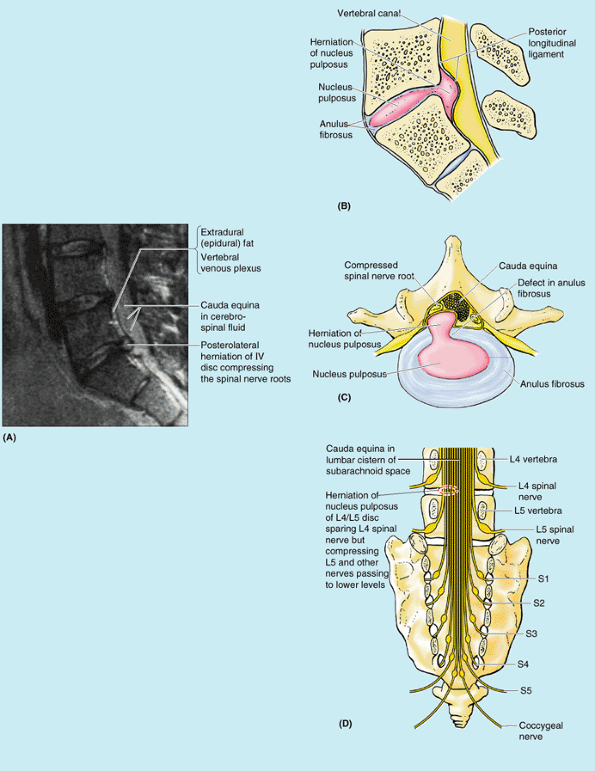 |
|
Figure B4.10. Herniation of nucleus pulposus. A. Left medial view, median MRI of lumbosacral region. B. Medial view, right half of hemisected lumbosacral joint. C. Superior view, transversely-sectioned herniated IV disc. D. Posterior view, cauda equina.
|
flexing of the vertebral column. The general rule is that when an IV
disc protrudes, it usually compresses the nerve root numbered one
inferior to the disc; for example, the L5 nerve is compressed by an
L4–L5 IV disc herniation and the S1 nerve by a L5–S1 IV disc
herniation. Recall that in the thoracic and lumbar regions the IV disc
forms the inferior half of the anterior border of the IV foramen and
that the superior half is formed by the bone of the body of the
superior vertebra (Fig. 4.8). The spinal nerve
roots descend to the IV foramen from which the spinal nerve formed by
their merging will exit. The nerve that exits a given IV foramen passes
through the superior bony half of the foramen and thus lies above and
is not affected by a herniating disc at that level. However, the nerve
roots passing to the IV foramen immediately and farther below pass
directly across the area of herniation (i.e., herniation of the L4–L5
disc affects the L5 nerve root). Symptom-producing IV disc protrusions
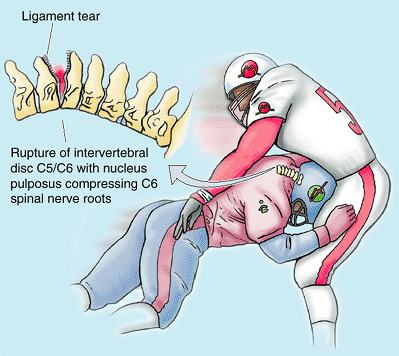
occur in the cervical region almost as often as in the lumbar region.
as might occur during a head-on collision or during illegal head
blocking in football, for example, may rupture the IV disc posteriorly
without fracturing the vertebral body (Fig. B4.11).
In this region, the IV discs are centrally placed in the anterior
border of the IV foramen, and a herniating disc compresses the nerve
actually exiting at that level (rather than the level below as in the
lumbar region). However, recall that cervical spinal nerves exit
superior to the vertebra of the same number, so the numerical
relationship of herniating disc to nerve affected is the same (e.g.,
the cervical IV discs most commonly ruptured are those between C5–C6
and C6–C7, compressing spinal nerve roots C6 and C7, respectively). IV
disc protrusions result in pain in the neck, shoulder, arm, and hand.
Any sport in which movement causes downward or twisting pressure on the
neck or lower back may produce herniation of the nucleus pulposus. The
most common sports involved are bowling, tennis, jogging, football,
hockey, weight lifting, and gymnastics.
 |
|
Figure B4.11
|
The capsule is attached to the margins of the articular surfaces of the
articular processes of adjacent vertebrae. Accessory ligaments unite
the laminae, transverse processes, and spinous processes and help
stabilize the joints.
between the articular processes; the shape and disposition of the
articular surfaces determine the types of movement possible. The range
(amount) of movement is determined by the size of the IV disc relative
to that of the vertebral body. In the cervical and lumbar regions,
these joints bear some weight, sharing this function with the IV discs
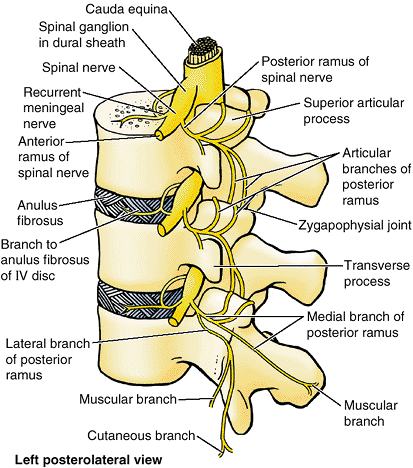
particularly during lateral flexion. The zygapophysial joints are
innervated by articular branches that arise from the medial branches of
the posterior rami of spinal nerves (Fig. 4.13).
As these nerves pass posteroinferiorly, they lie in grooves on the
posterior surfaces of the medial parts of the transverse processes.
Each articular branch supplies two adjacent joints; therefore, each
joint is supplied by two nerves.
because they are close to the IV foramina through which the spinal
nerves emerge from the vertebral canal. When these joints are injured
or develop osteophytes during aging (osteoarthritis), the related spinal nerves are often affected. This causes pain along the distribution patterns of the dermatomes and spasm in the muscles derived from the associated myotomes (a myotome consists all the muscles or parts of muscles receiving innervation from one spinal nerve).
is a procedure historically used for treatment of back pain caused by
disease of these joints. The nerves are sectioned near the joints or
are destroyed by radiofrequency percutaneous rhizolysis (G. rhiza, root + G. lysis,
dissolution). The denervation is directed at the articular branches of
two adjacent posterior rami of the spinal nerves because each joint
receives innervation from both the nerve exiting that level and the
superjacent nerve.
 |
|
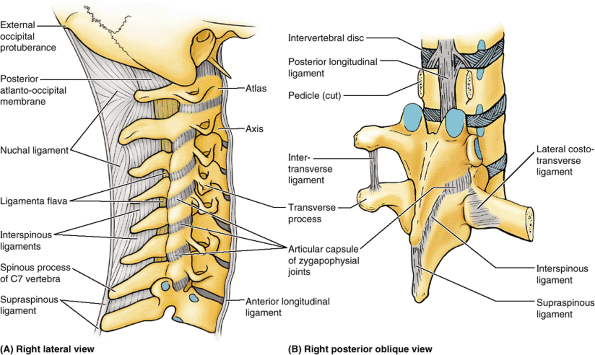
Figure 4.12. Joints and ligaments of vertebral column. A.
The ligaments in the cervical region are shown. Superior to the prominent spinous process of C7 (vertebra prominens), the spinous processes are deeply placed and attached to an overlying nuchal ligament. B. The ligaments in the thoracic region are shown. The pedicles of the superior two vertebrae have been sawn through and the vertebral arches removed to reveal the posterior longitudinal ligament. Intertransverse, supraspinous, and interspinous ligaments are demonstrated in association with the vertebrae with intact vertebral arches. |
yellow). These yellow ligaments extend almost vertically from the
lamina above to the lamina below, those of opposite sides meeting and
blending in the midline (Fig. 4.11). The
ligaments bind the lamina of the adjoining vertebrae together, forming
alternating sections of the posterior wall of the vertebral canal. The
ligamenta flava are long, thin, and broad in the cervical region,
thicker in the thoracic region, and thickest in the lumbar region.
These ligaments resist separation of the vertebral lamina by arresting
abrupt flexion of the vertebral column and thereby preventing injury to
the IV discs. The strong elastic ligamenta flava help preserve the
normal curvatures of the vertebral column and assist with straightening
of the column after flexing.
The thin interspinous ligaments connect adjoining spinous processes,
attaching from the root to the apex of each process. The cord-like supraspinous ligament, which connects the apices (tips) of the spinous processes from C7 to the sacrum, merges superiorly with the nuchal ligament (L. ligamentum nuchae), the strong, broad, median band at the back of the neck (Fr. nuque, back of neck) (Fig. 4.12A).
Unlike the interspinous and supraspinous ligaments, the nuchal ligament
is composed of thickened fibroelastic tissue, extending from the
external occipital protuberance and posterior border of the foramen
magnum to the spinous processes of the cervical vertebrae. Because of
the shortness of the C3–C5 spinous processes, the nuchal ligament
substitutes for bone in providing muscular
attachments. The intertransverse ligaments,
connecting adjacent transverse processes, consist of scattered fibers
in the cervical region and fibrous cords in the thoracic region (Fig. 4.12B). The intertransverse ligaments in the lumbar region are thin and membranous.
 |
|
Figure 4.13. Innervation of zygapophysial joints.
The posterior rami arise from the spinal nerves outside the IV foramen and divide into medial and lateral branches; the medial branch gives rise to articular branches distributed to the zygapophysial joint at that level and to the joint one level inferior to its exit. Thus, each zygapophysial joint receives articular rami from the medial branch of the posterior rami of two adjacent spinal nerves; the medial branches of both posterior rami would have to be ablated to denervate a zygapophysial joint. |
and zygapophysial joints. The relative thickness of the discs
determines the degree of mobility. The disposition of the zygapophysial
joints controls the type of movement between adjacent vertebrae. The
anterior longitudinal ligament resists hyperextension; all other
ligaments resist forms of flexion.
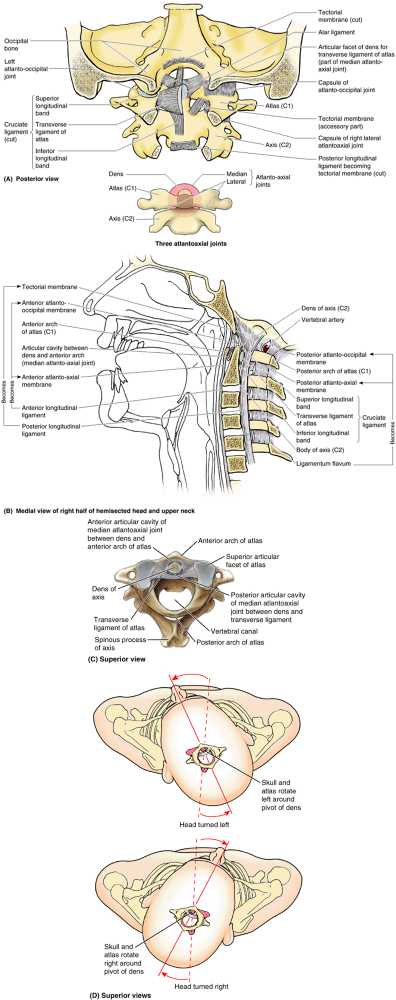
atlanto-occipital joints. formed between the atlas (C1 vertebra) and
the occipital bone of the cranium, and the atlantoaxial joints, formed
between the atlas and axis (C2 vertebra) (Fig. 4.14). The Greek word atlanto
refers to the atlas. The craniovertebral joints are synovial joints
that have no IV discs. Their design gives a wider range of movement
than in the rest of the vertebral column. The articulations involve the
occipital condyles, atlas, and axis.
the atlanto-occipital joints, permit nodding of the head, such as the
neck flexion and extension occurring when indicating approval (the
“yes” movement). These joints also permit sideways tilting of the head.
The main movement is flexion, with a little lateral bending and
rotation. They are synovial joints of the condyloid type and have thin,
loose articular capsules. The cranium and C1 are also connected by anterior and posterior atlanto-occipital membranes, which extend from the anterior and posterior arches of C1 to the anterior and posterior margins of the foramen magnum (Figs. 4.14B and 4.15).
The anterior membranes are composed of broad, densely woven fibers
(especially centrally where they are continuous with the anterior
longitudinal ligament); the posterior membranes are broad but
relatively weak. The atlanto-occipital membranes help prevent excessive
movement of these joints.
(between the dens of C2 and the anterior arch of the atlas). The
lateral atlantoaxial joints are gliding-type synovial joints, whereas
the median atlantoaxial joint is a pivot joint. Movement at all three
atlantoaxial joints permits the head to be turned from side to side (Fig. 4.14D),
as occurs when rotating the head to indicate disapproval (the “no”
movement). During this movement, the cranium and C1 rotate on C2 as a
unit. During rotation of the head, the dens of C2 is the axis or pivot
that is held in a socket or collar formed anteriorly by the anterior
arch of the atlas and posteriorly by the transverse ligament of the atlas (Fig. 4.14A–C),
a strong band extending between the tubercles on the medial aspects of
the lateral masses of C1 vertebrae. Vertically oriented but much weaker
superior and inferior longitudinal bands pass from the trans
verse ligament to the occipital bone superiorly and to the body of C2
inferiorly. Together, the transverse ligament and the longitudinal
bands form the cruciate ligament (formerly the cruciform ligament), so named because of its resemblance to a cross.
 |
|
Figure 4.14. Craniovertebral joints and ligaments. A.
Ligaments of the atlanto-occipital and atlantoaxial joints. The tectorial membrane and the right side of the cruciate ligament of the atlas have been removed to show the attachment of the right alar ligament to the dens of vertebra C2 (axis). B. The hemisected craniovertebral region shows the median joints and membranous continuities of the ligamenta flava and longitudinal ligaments in the craniovertebral region. C. The articulated atlas and axis showing that the median atlantoaxial joint is formed as the anterior arch and the transverse ligament of the atlas form a socket for the dens of the axis. D. During rotation of head, the cranium and atlas rotate as a unit around the pivot of the dens when the head is turned side to side (the “no” movement). |
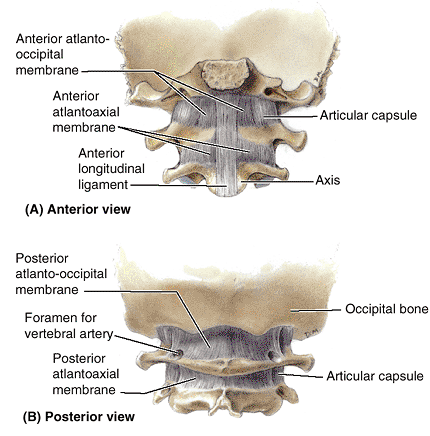 |
|
Figure 4.15. Membranes of craniovertebral joints. A.
Only the thicker, most anterior part of the anterior longitudinal ligament is included here to demonstrate its superior continuation as the anterior atlantoaxial membrane and anterior atlanto-occipital membrane. Laterally, the membranes blend with the articular capsules of the lateral atlantoaxial and atlanto-occipital joints. B. The posterior atlanto-occipital and atlantoaxial membranes span the gaps between the posterior arch of the atlas (C1) and the occipital bone (posterior margin of the foramen magnum) superiorly and the laminae of the axis (C2) inferiorly. The vertebral arteries penetrate the atlanto-occipital membrane before traversing the foramen magnum. |
These short, rounded cords, approximately 0.5 cm in diameter (just
smaller than a pencil), attach the cranium to vertebra C1 and serve as
check ligaments, preventing excessive rotation at the joints.
is the strong superior continuation of the posterior longitudinal
ligament across the median atlantoaxial joint through the foramen
magnum to the central floor of the cranial cavity. It runs from the
body of C2 to the internal surface of the occipital bone and covers the
alar and transverse ligaments.
movement of the head. The atlantoaxial joints enable the “no”
(rotational) movement of the head. Alar ligaments limit rotation.
the dens of the C2 vertebra. Fractures of the dens make up about 40% of
axis fractures. The most common dens fracture occurs at its base—that
is, at its junction with the body of the axis (Fig. B4.12A).
Often these fractures are unstable (do not reunite) because the
transverse ligament of the atlas becomes interposed between fragments (Crockard et al., 1993)
and because the separated fragment (the dens) no longer has a blood
supply, resulting in avascular necrosis. Almost as common are fractures
of the vertebral body inferior to the base of the dens (Fig. B4.12B–E).
This type of fracture heals more readily because the fragments retain
their blood supply. Other dens fractures result from abnormal
ossification patterns.
Pathological softening of the transverse and adjacent ligaments,
usually resulting from disorders of connective tissue, may also cause
atlantoaxial subluxation (Bogduk and Macintosh, 1984);
20% of people with Down syndrome exhibit laxity or agenesis of this
ligament. Dislocation owing to transverse ligament rupture or agenesis
is more likely to cause spinal cord compression than that resulting
from fracture of the dens (Fig. B4.13B).
In fractures of the dens, the dens fragment is held in place against
the anterior arch of the atlas by the transverse ligament, and the dens
and atlas move as a unit. In the absence of a competent ligament, the
upper cervical region of the spinal cord may be compressed between the
approximated posterior arch of the atlas and the dens (Fig. B4.13A), causing paralysis of all four limbs (quadriplegia),
or into the medulla of the brainstem, resulting in death. Approximately
one third of the atlas ring is occupied by the dens, one third by the
spinal cord, and the remaining third by the fluid-filled space and
tissues surrounding the cord: Steele Rule of Thirds (Fig. B4.13C & D).
This explains why some patients with anterior displacement of the atlas
may be relatively asymptomatic until a large degree of movement
(greater than one third of the diameter of the atlas ring) occurs.
Sometimes inflammation in the craniovertebral area may produce
softening of the ligaments of the craniovertebral joints and cause
dislocation of the atlantoaxial joints. Sudden movement of a patient
from a bed to a chair, for example, may produce posterior displacement
of the dens and injury to the spinal cord.
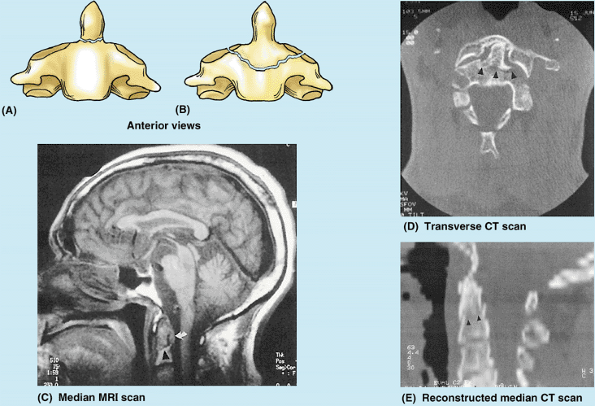 |
|
Figure B4.12. Fractures of dens. Black arrowheads (in C, D, & E), fracture lines at root or base of dens; white arrow (in C), transverse ligament of atlas.
|
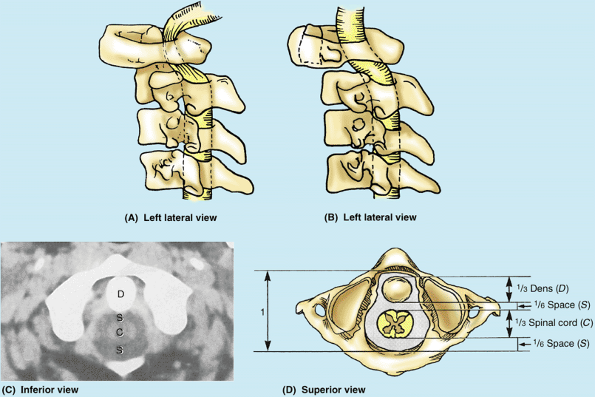 |
|
Figure B4.13. Rupture of transverse ligament of atlas. A.
This left lateral view demonstrates that subluxation of the median atlantoaxial joint is allowed by rupture of the transverse ligament of the atlas. The atlas moves, but the dens is fixed. B. This left lateral view of a fracture of the dens shows that the dens and atlas move together as a unit because the transverse ligament holds the dens to the anterior arch of the atlas. C and D. Inferior view transverse CT scan and interpretive drawing showing a normal median atlantoaxial joint and demonstrating Steele’s Rule of Thirds |
ligament of the atlas. Consequently, combined flexion and rotation of
the head may tear one or both alar ligaments. Rupture of an alar
ligament results in an increase of approximately 30% in the range of
movement to the contralateral side (Dvorak et al., 1988).
the C2 spinal nerve. When the neck is severely hyperextended while the
head is turned to the side, the spinal ganglion of the C2 nerve on the
opposite side may be compressed between C1 and C2 vertebrae. This may
be followed by prolonged severe headaches and excruciating
cervico-occipital pain.
according to the region and the individual. Some people are capable of
extraordinary movements, such as acrobats who began their training
during early childhood. The normal range of movement possible in
healthy young adults is typically reduced by 50% or more as they age.
from the compressibility and elasticity of the IV discs. The vertebral
column is capable of flexion, extension, lateral flexion and extension,
and rotation (torsion) (Fig. 4.16). Bending of the vertebral column to the right or left from the neutral (erect) position is lateral flexion (bending); returning to the erect posture from a position of lateral flexion is lateral extension.
-
Thickness, elasticity, and compressibility of the IV discs.
-
Shape and orientation of the zygapophysial joints.
-
Tension of the joint capsules of the zygapophysial joints.
-
Resistance of the back muscles and ligaments (e.g., the ligamenta flava and the posterior longitudinal ligament).
-
Attachment to the thoracic (rib) cage.
-
Bulk of surrounding tissue.
column are discussed later in this chapter. Movements are not produced
exclusively, however, by the back muscles. They are assisted by gravity
and the action of the anterolateral abdominal muscles (see Chapter 2).
resilient nuclei pulposi of the IV discs (serving as the axis of
movement) and at the zygapophysial joints. The orientation of the
latter joints permits some movements and restricts others. With the
exception perhaps of C1–C2, movement never occurs at a single segment
of the column. Although movements between adjacent vertebrae are
relatively small, especially in the thoracic region, the summation of
all the small movements produces a considerable range of movement of
the vertebral column as a whole (e.g., when flexing to touch the toes).
Movements of the vertebral column are freer in the cervical and lumbar
regions than elsewhere. Flexion, extension, lateral flexion, and
rotation of the neck are especially free because the:
-
IV discs, although thin relative to most other discs, are thick relative to the size of the vertebral bodies at this level.
-
Articular surfaces of the zygapophysial joints are relatively large and the joint planes are almost horizontal.
-
Joint capsules of the zygapophysial joints are loose.
-
Neck is relatively slender (with less surrounding soft tissue bulk compared with the trunk).
The sagittally oriented joint planes of the lumbar region are conducive
to flexion and extension. Extension of the vertebral column is most
marked in the lumbar region and is usually more extensive than flexion;
however, the interlocking articular processes here prevent rotation (Table 4.3).
The lumbar region, like the cervical region, has IV discs that are
large (the largest discs occur here) relative to the size of the
vertebral bodies. Lateral flexion of the vertebral column is greatest
in the cervical and lumbar regions (Fig. 4.16B).
thin relative to the size of the vertebral bodies. Relative stability
is also conferred on this part of the vertebral column, through its
connection to the sternum by the ribs and costal cartilages. The joint
planes here lie on an arc that is centered on the vertebral body,
permitting rotation in the thoracic region (Fig. 4.16C).
This rotation of the upper trunk, in combination with the rotation
permitted in the cervical region and that at the atlantoaxial joints,
enables the torsion of the axial skeleton that occurs as one looks back
over the shoulder. However, flexion is limited in the thoracic region,
including lateral flexion.
 |
|
Figure 4.16. Movements of vertebral column. A.
Flexion (forward bending) and extension (backward bending), both in the median plane, are shown. Flexion and extension are occurring primarily in the cervical and lumbar regions. B. Lateral flexion (to the right or left in a frontal plane) is shown, also occurring mostly in the cervical and lumbar regions. C. Rotation around a longitudinal axis, which occurs primarily at the craniovertebral joints (augmented by the cervical region) and the thoracic region, is shown. |
permits a considerable amount of movement as well as support and
protection, excessive or sudden violent movement or movement of a type
not permitted in a specific region, is likely to result in fractures,
dislocations, and fracture–dislocations of the vertebral column. Sudden
forceful flexion, as occurs in automobile accidents or from a violent
blow to the back of the head, commonly produces a crush or compression fracture
of the body of one or more vertebrae. If violent anterior movement of
the vertebra occurs in combination with compression, a vertebra may be
displaced anteriorly on the vertebra inferior to it (e.g., dislocation
of C6 or C7 vertebrae) (see clinical correlation [blue] box “Dislocation of the Cervical Vertebrae,”
in this chapter). Usually this displacement dislocates and fractures
the articular facets between the two vertebrae and ruptures the
interspinous ligaments. Irreparable injuries to the spinal cord
accompany most severe flexion injuries of the vertebral column.
the vertebral column and spinal cord. Head butting or illegal face
blocking in football may lead to a hyperextension injury of the neck (Fig. B4.14A).
Such severe hyperextension is most likely to injure posterior parts of
the vertebrae, fracturing by crush or compression of the vertebral
arches and their processes. Fractures of cervical vertebrae may radiate
pain to the back of the neck and scapular region because the same
spinal sensory ganglia and spinal cord segments receiving pain impulses
from the vertebrae are also involved in supplying neck muscles.
Hyperextension (“whiplash” injury) of the neck also occurs during
rear-end motor vehicle collisions (Fig. B4.14B),
an association especially well known to litigation lawyers. In these
types of hyperextension injuries, the anterior longitudinal ligament is
severely stretched and may be torn.
occur as the head “rebounds” after the hyperextension, snapping the
head forward onto the thorax. “Facet jumping” or locking of the
cervical vertebrae may occur because of dislocation of the vertebral
arches (see clinical correlation (blue) box “Dislocation of the Cervical Vertebrae,”
in this chapter). Severe hyperextension of the head on the upper neck
may, in addition to producing a cervical spondylolysis or hangman’s
fracture (see clinical correlation (blue) box “Fracture and Dislocation of the Axis,”
in this chapter), rupture the anterior longitudinal ligament and the
adjacent anulus fibrosus of the C2–C3 IV disc. If this occurs, the
cranium, C1, and the anterior portion (dens and body) of C2 are
separated from the rest of the axial skeleton (Fig. B4.14C),
and the spinal cord is usually severed. Persons with this severe injury
seldom survive. Football, diving, falls from horses, and motor vehicle
collisions cause most fractures of the cervical region of the vertebral
column. Symptoms range from vague aches to progressive loss of motor
and sensory functions.
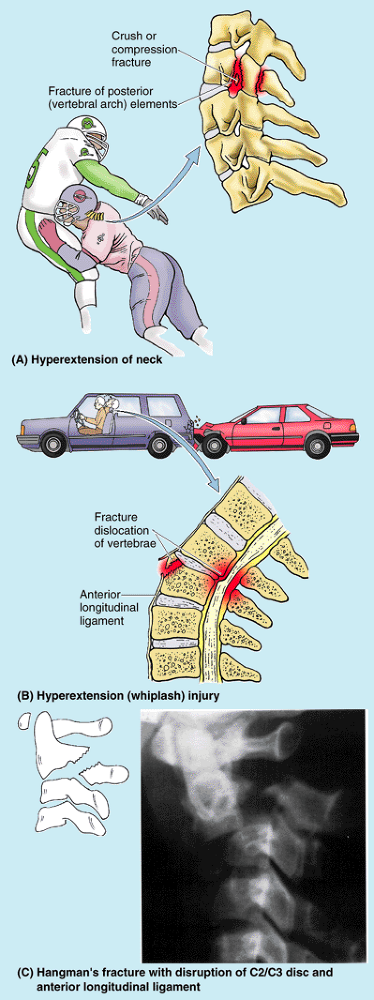 |
|
Figure B4.14
|
regions is uncommon because of the interlocking of their articular
processes. The transition from the relatively inflexible thoracic
region to the much more mobile lumbar region occurs abruptly,
unfortunately. Consequently, vertebrae T11 and especially T12 (which
participates in rotatory movements superiorly but only flexion and
extension movements inferiorly) are the most commonly fractured
noncervical vertebrae (i.e., a “broken back” vs. a “broken neck”).
Most agree that spondylolysis of L5, or susceptibility to it, probably
results from a failure of the centrum of L5 to unite adequately with
the neural arches at the neurocentral joint during development (see “Ossification of Vertebrae,” earlier in this chapter). The anatomical basis of spondylolisthesis was discussed in Chapter 3
in relation to impingement on the pelvic inlet. Spondylolisthesis at
the L5–S1 articulation may (but does not necessarily) result in
pressure on the spinal nerves of the cauda equina as they pass into the
superior part of the sacrum, causing back and lower limb pain.
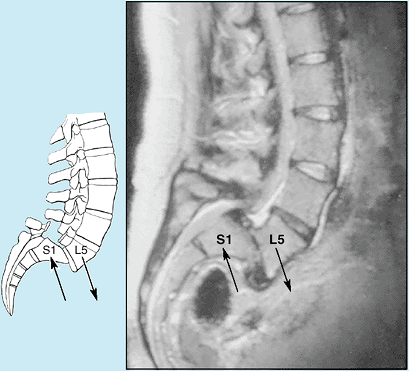 |
|
Figure B4.15. Spondylolisthesis secondary to spondylolysis.
|
consequently most vulnerable to injury). Flexion and extension occur
primarily in the cervical and lumbar regions, whereas rotation occurs
in the cervical and thoracic regions.
noting that the primary curvatures are in the same direction as the
main curvatures of the fetal vertebral column. The primary curvatures
are retained throughout life as a consequence of differences in height
between the anterior and posterior parts of the vertebrae.
that result from extension from the flexed fetal position. They begin
to appear during the fetal period but do not become obvious until
infancy. Secondary curvatures are maintained primarily by differences
in thickness between the anterior and the posterior parts of the IV
discs. The cervical curvature becomes
fully evident when an infant begins to raise (extend) its head while
prone and to hold its head erect while sitting. The lumbar curvature
becomes apparent when an infant begins to assume the upright posture,
standing and walking. This curvature, generally more pronounced in
females, ends at the lumbosacral angle formed at the junction of L5 vertebra with the sacrum (Fig. 4.1D). The sacral curvature also differs in males and females, that of the female reduced so that the coccyx protrudes less into the pelvic outlet (see Chapter 3).
(shock-absorbing resilience) to the vertebral column, further
augmenting that provided by the IV discs. When the load borne by the
vertebral column is markedly increased (as by carrying a heavy backpack
or another person on one’s shoulders), both the IV discs and the
flexible curvatures are compressed (i.e., the curvatures tend to
increase). While the flexibility provided by the IV discs is passive
and limited primarily by the zygapophysial joints and longitudinal
ligaments, that provided by the curvatures is actively (dynamically)
resisted by the contraction of
muscle
groups antagonistic to the movement (e.g., the long extensors of the
back resist increased thoracic kyphosis, whereas the abdominal flexors
resist increased lumbar lordosis). Carrying additional weight anterior
to the body’s normal gravitational axis (e.g., abnormally large
breasts, a pendulous abdomen, or carrying a young child) also tends to
increase these curvatures. The muscles that provide resistance to the
increase in curvature often ache when the weight is borne for extended
periods.
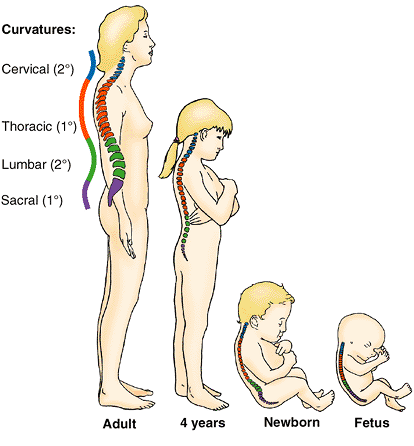 |
|
Figure 4.17. Curvatures of vertebral column.
The four curvatures of the adult vertebral column—cervical, thoracic, lumbar, and sacral—are contrasted with the C-shaped curvature of the vertebral column during fetal life, when only the primary (1°) curvatures exist. The secondary (2°) curvatures develop during infancy and childhood. |
for long periods of time, one usually “cycles” between back flexion
(slumping) and extension (sitting up straight) to minimize stiffness
and fatigue (Fig. 4.18). This allows
alternation between the active support provided by the extensor muscles
of the back and the passive resistance to flexion provided by ligaments.
developmental; secondary (cervical and lumbar) curvatures are acquired
in relation to erect human posture. The curvatures provide
shock-absorbing resilience and flexibility to the axial skeleton.
Extensors of the back and abdominal flexors provide dynamic support to
maintain the curvatures.
 |
|
Figure 4.18. Flexion and extension while sitting. During prolonged sitting (especially when no back support is provided), flexion (A) and extension (B) occur alternately, transferring the load between muscles (active) and ligaments (passive).
|
have an individual stand in the anatomical position. Inspect the
profile of the vertebral column from the person’s side (Fig. B4.16A–C) and then from the posterior aspect (Fig. B4.16D).
With the person bending over, observe the ability to flex directly
forward and whether the back is level once the flexed position is
assumed (Fig. B4.16E).
Abnormal curvatures in some people result from developmental anomalies;
in others, the curvatures result from pathological processes. The most
prevalent metabolic disease of bone occurring in the elderly,
especially in women, is osteoporosis. It
is characterized by net demineralization of the bones caused by a
disruption of the normal balance of calcium deposition and resorption.
As a result, the quality of bone is reduced and atrophy of skeletal
tissue occurs. Although osteoporosis affects the entire skeleton, the
most affected areas are the neck of the femur (thigh bone), the
vertebrae, the metacarpals (bones of the hand), and the radius (of the
forearm). These bones become weakened and brittle, and are subject to
fracture.
is a colloquial name for excess thoracic kyphosis in older women
resulting from osteoporosis. However, excess kyphosis also occurs in
elderly men (Swartz, 2002). Osteoporosis especially affects the horizontal trabeculae of the trabecular (spongy) bone of the vertebral body (Fig. 4.3).
The remaining, unsupported vertical trabeculae are less able to resist
compression and sustain compression fractures, resulting in short and
wedge-shaped thoracic vertebrae (Fig. B4.8A).
Progressive erosion and collapse of vertebrae also result in an overall
loss of height, and the excess kyphosis leads to an increase in the AP
diameter of the thorax.
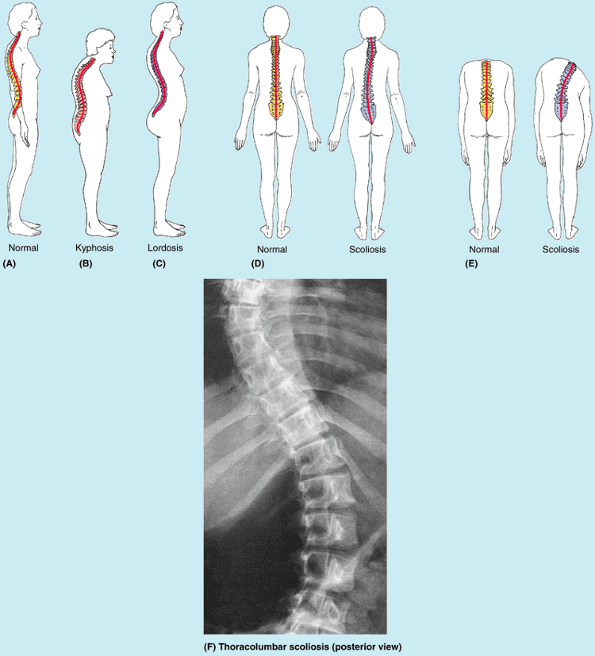 |
|
Figure B4.16
|
is characterized by an anterior rotation of the pelvis (the upper
sacrum tilts anteroinferiorly) at the hip joints, producing an abnormal
increase in the lumbar curvature; the vertebral column curves more
anteriorly (Fig. B4.16C). This abnormal extension deformity
is often associated with weakened trunk musculature, especially the
anterolateral abdominal muscles. To compensate for alterations to their
normal line of gravity, women develop a temporary excess lumbar
lordosis during late pregnancy. This lordotic curvature may cause low
back pain, but the discomfort normally disappears soon after
childbirth. Obesity in both sexes can also cause excess lumbar lordosis
and low back pain because of the increased weight of the abdominal
contents (e.g., “potbelly”) anterior to the normal line of gravity.
Loss of weight and exercise of the anterolateral abdominal muscles
facilitate correction of this type of excess lordosis.
The spinous processes turn toward the cavity of the abnormal curvature,
and when the individual bends over, the ribs rotate posteriorly
(protrude) on the side of the increased convexity. Scoliosis is the
most common deformity of the vertebral column in pubertal girls (ages
12–15 years). Asymmetrical weakness of the intrinsic back muscles (myopathic scoliosis), failure of half of a vertebra to develop (hemivertebra),
and a difference in the length of the lower limbs are causes of
scoliosis. If the lengths of the lower limbs are not equal, a
compensatory pelvic tilt may lead to a functional static scoliosis.
When a person is standing, an obvious inclination or listing to one
side may be a sign of scoliosis that is secondary to a herniated IV
disc. Habit scoliosis is supposedly caused
by habitual standing or sitting in an improper position. When the
scoliosis is entirely postural, it disappears during maximum flexion of
the vertebral column. Sometimes there is kyphoscoliosis,
excess thoracic kyphosis combined with scoliosis, in which an abnormal
AP diameter produces a severe restriction of the thorax and lung
expansion (Swartz, 2002).
Parent arteries of periosteal, equatorial, and spinal branches occur at
all levels of the vertebral column, in close association with it, and
include the following arteries (described in detail in other chapters):
-
Vertebral and ascending cervical arteries in the neck (Chapter 8).
-
The major segmental arteries of the trunk:
-
Posterior intercostal arteries in the thoracic region (Chapter 1).
-
Subcostal and lumbar arteries in the abdomen (Chapter 2).
-
Iliolumbar and lateral and medial sacral arteries in the pelvis (Chapter 3).
arise from these arteries as they cross the external (anterolateral)
surfaces of the vertebrae. Spinal branches enter the IV foramina and
divide. Smaller anterior and posterior vertebral canal branches
pass to the vertebral body and vertebral arch, respectively, and give
rise to ascending and descending branches that anastomose with the
spinal canal branches of adjacent levels. Anterior vertebral canal
branches send nutrient arteries anteriorly into the vertebral bodies that supply most of the red marrow of the central vertebral body (Bogduk, 1997). The
larger branches of the spinal branches continue as terminal radicular or segmental medullary arteries
distributed to the posterior and anterior roots of the spinal nerves
and their coverings and to the spinal cord, respectively (see “Vasculature of the Spinal Cord and Spinal Nerve Roots,” later in this chapter).
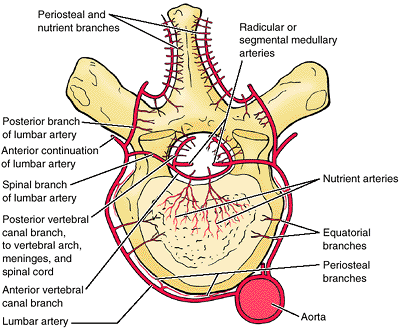 |
|
Figure 4.19. Blood supply of vertebrae.
Typical vertebrae are supplied by segmental arteries—here lumbar arteries. In the thoracic and lumbar regions, each vertebra is encircled on three sides by paired intercostal or lumbar arteries that arise from the aorta. The segmental arteries supply equatorial branches to the vertebral body, and posterior branches supply the vertebral arch structures and the back muscles. Spinal branches enter the vertebral canal through the IV foramina to supply the bones, periosteum, ligaments, and meninges that bound the epidural space and radicular or segmental medullary arteries that supply nervous tissue (spinal nerve roots and spinal cord). |
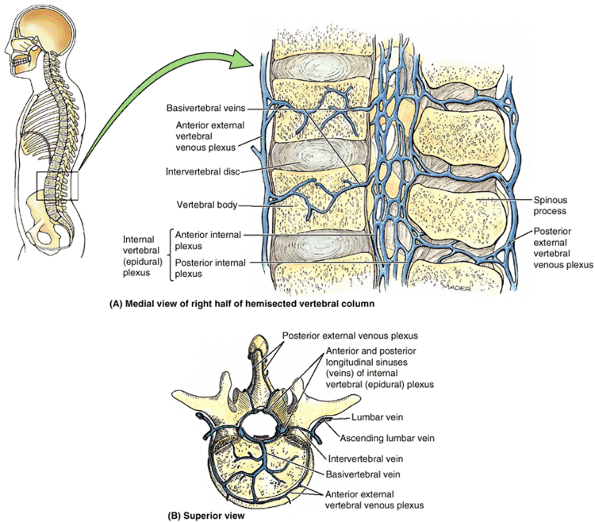 |
|
Figure 4.20. Venous drainage of vertebral column. A.
The venous drainage parallels the arterial supply and enters the external and internal vertebral venous plexuses. There is also anterolateral drainage from the external aspects of the vertebrae into segmental veins. B. The superior view of a lumbar vertebra is shown. The vertebral canal contains a dense plexus of thin-walled valveless veins, the internal vertebral venous plexuses, which surround the dura mater. Anterior and posterior longitudinal venous sinuses can be identified in the internal vertebral venous plexus. Basivertebral veins from the vertebral body drain primarily into the anterior internal vertebral venous plexus, but they may also drain to the anterior external plexus. |
These plexuses communicate through the intervertebral foramina. Both
plexuses are densest anteriorly and posteriorly and relatively sparse
laterally. The large, tortuous basivertebral veins
form within the vertebral bodies. They emerge from foramina on the
surfaces of the vertebral bodies (mostly the posterior aspect) and
drain into the anterior external and especially the anterior internal
vertebral venous plexuses, which may form large longitudinal sinuses.
The intervertebral veins receive veins from the spinal cord and vertebral venous plexuses as they accompany the spinal
nerves through the IV foramina to drain into the vertebral veins of the neck and segmental (intercostal, lumbar, and sacral) veins of the trunk.
arteries supply the vertebral column. Internal and external vertebral
venous plexuses collect blood from the vertebrae and drain, in turn,
into the vertebral veins of the neck and the segmental veins of the
trunk.
described with these joints), the vertebral column is innervated by (recurrent) meningeal branches of the spinal nerves (Fig. 4.21).
These rarely described or depicted branches are the only branches to
arise from the mixed spinal nerve, arising immediately after it is
formed and before its division into anterior and posterior rami or from
the anterior ramus immediately after its formation. Two to four of
these fine branches arise on each side at all vertebral levels. Close
to their origin, the meningeal branches receive communicating branches
from the nearby gray rami communicantes. As the spinal nerves exit the
IV foramina, most of the meningeal branches run back through the
foramina into the vertebral canal (hence the alternate term recurrent).
However, some branches remain outside the canal and are distributed to
the anterolateral aspect of the vertebral bodies and IV discs. They
supply the periosteum and especially the anuli fibrosi and anterior
longitudinal ligament. Inside the vertebral canal, transverse,
ascending, and descending branches distribute nerve fibers to the:
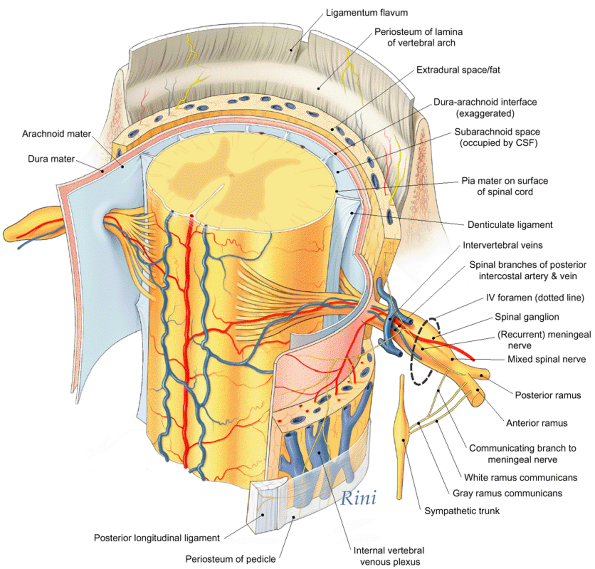 |
|
Figure 4.21. Innervation of periosteum and ligaments of vertebral column and of meninges.
Except for the zygapophysial joints and external elements of the vertebral arch, the fibroskeletal structures of the vertebral column (and the meninges) are supplied by the (recurrent) meningeal nerves. Although usually omitted from diagrams and illustrations of spinal nerves, these fine nerves are the first branches to arise from all 31 pairs of spinal nerves and are the nerves that initially convey localized pain sensation from the back produced by acute herniation of an IV disc or from sprains, contusions, fractures, or tumors of the vertebral column itself. (Based on Frick H, Kummer B, Putz R: Wolf-Heidegger’s Atlas of Human Anatomy, 4th ed. Basel, Karger AG, 1990:476.
) |
-
Periosteum (covering the surface of the posterior vertebral bodies, pedicles, and laminae).
-
Ligamenta flava.
-
Anuli fibrosi of the posterior and posterolateral aspect of the IV discs.
-
Posterior longitudinal ligament.
-
Spinal dura mater.
-
Blood vessels within the vertebral canal.
ligaments supply pain receptors; those to the anuli fibrosi and
ligaments also supply receptors for proprioception (the sense of one’s
position). Sympathetic fibers to the blood vessels stimulate
vasoconstriction. Innervation of the dura mater is discussed later in
this chapter.
of adjacent posterior rami. (Recurrent) meningeal branches of spinal
nerves supply most bone (periosteum), IV discs, and ligaments as well
as the coverings of the spinal cord. These two (groups of) nerves
convey all localized pain from the vertebral column.
is an immense health problem, second only to the common cold as a
reason people visit their doctors. In terms of health factors causing
lost work days, backache is second only to headache. Annual costs for
treatment of back pain are staggering. Yet the anatomical bases for the
pain, especially the nerves initially involved in sensing and carrying
pain from the vertebral column itself, are rarely described.
-
Fibroskeletal structures: periosteum, ligaments, and the anuli fibrosi of IV discs.
-
Meninges: the coverings of the spinal cord.
-
Synovial joints: the capsules of the zygapophysial joints.
-
Muscles: the intrinsic muscles of the back.
-
Nervous tissue: spinal nerves or nerve roots exiting the IV foramina.
meningeal branches of the spinal nerves and the next two are innervated
by posterior rami (articular and muscular branches). Pain resulting
from nervous tissue—that is, caused by compression or irritation of
spinal nerves or nerve roots—is referred to the area (dermatome)
supplied by that nerve (see clinical correlation [blue] box “Herniation of the Nucleus Pulposus,”
in this chapter) but may be accompanied by localized pain. Pain related
to the meninges is relatively rare and is discussed later in this
chapter; it is generally not considered to be a factor in back pain.
is generally associated with aging (osteoarthritis) or disease
(rheumatoid arthritis) of the joints. Pain from vertebral fractures and
dislocations is no different from that from other bones and joints: The
sharp pain following a fracture is mostly periosteal in origin, whereas
pain from dislocations is ligamentous. The acute localized pain
associated with an IV disc herniation undoubtedly emanates from the
disrupted posterolateral anulus fibrosis and impingement on the
posterior longitudinal ligament. Pain in all of these latter instances
is conveyed initially by the meningeal branches of the spinal nerves.
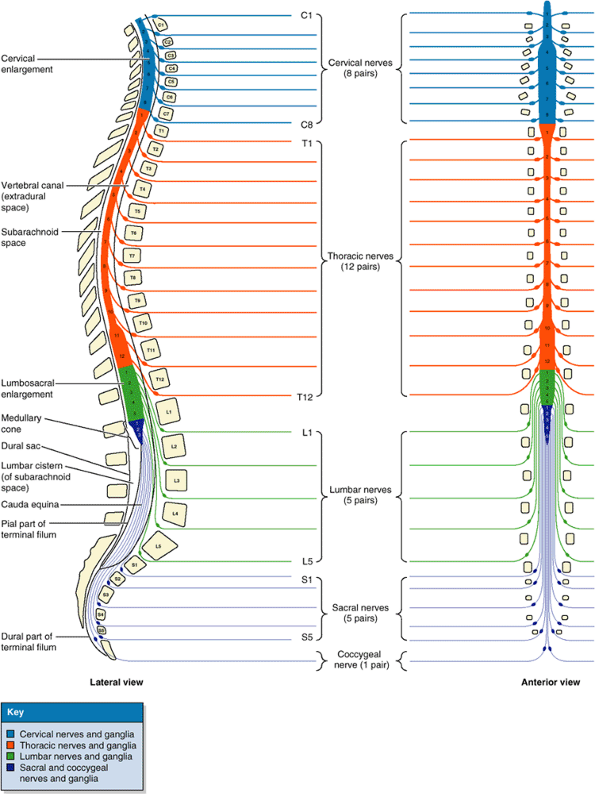 |
|
Figure 4.22. Vertebral column, spinal cord, spinal ganglia, and spinal nerves.
Lateral and posterior views illustrating the relation of the spinal cord segments (the numbered segments) and spinal nerves to the adult vertebral column. |
and the neurovascular structures that supply them are located within
the vertebral canal (Fig. 4.21).
reflex center and conduction pathway between the body and the brain.
This cylindrical structure, slightly flattened anteriorly and
posteriorly, is protected by the vertebrae and their associated
ligaments and muscles, the spinal meninges, and the cerebrospinal fluid
(CSF). The spinal cord begins as a continuation of the medulla oblongata (commonly called the medulla),
the caudal part of the brainstem. In adults, the spinal cord is 42–45
cm long and extends from the foramen magnum in the occipital bone to
the level of the L1 or L2 vertebra (Fig. 4.22). However, its tapering inferior end, the medullary cone,
may terminate as high as T12 vertebra or as low as L3 vertebra. Thus
the spinal cord occupies only the superior two thirds of the vertebral
canal.
extends from the C4 through T1 segments of the spinal cord, and most of
the anterior rami of the spinal nerves arising from it form the brachial plexus of nerves that innervates the upper limbs (see Chapter 6). The lumbosacral (lumbar) enlargement
extends from T11 through S1 segments of the spinal cord, inferior to
which the cord continues to diminish as the medullary cone. The
anterior rami of the spinal nerves arising from this enlargement make
up the lumbar and sacral plexuses of nerves that innervate the lower limbs (see Chapter 5).
roots are discussed in the Introduction. Readers are urged to read this
information now if they have not done so previously.
rootlets and roots that ultimately form one bilateral pair of spinal
nerves is designated a spinal cord segment,
the identity of which is the same as the spinal nerves arising from it.
Cervical spinal nerves (except C8) bear the same alphanumeric
designation as the vertebrae forming the inferior margin of the IV
foramina through which the nerve exits the vertebral canal, whereas the
more inferior spinal nerves bear the same alphanumeric designation as
the vertebrae forming the superior margin of their exit. The first
cervical nerves lack posterior roots in 50% of people, and the
coccygeal nerve may be absent altogether.
thus spinal cord segments lie approximately at the vertebral level of
the same number, the spinal nerves passing laterally to exit the
corresponding IV foramen. By the end of the embryonic period (8th
week), the tail-like caudal eminence has disappeared, and the number of
coccygeal vertebrae is reduced from six to four segments. The spinal
cord in the vertebral canal of the coccyx atrophies. During the fetal
period, the vertebral column grows faster than the spinal cord; as a
result, the cord “ascends” relative to the vertebral canal. At birth,
the tip of the medullary cone is at the L4–L5 level. Thus, in postnatal
life, the spinal cord is shorter than the vertebral column;
consequently, there is a progressive obliquity of the spinal nerve
roots (Figs. 4.22 and 4.23).
Because the distance between the origin of a nerve’s roots from the
spinal cord and the nerve’s exit from the vertebral canal increases as
the inferior end of the vertebral column is approached, the length of
the nerve roots also increases progressively. The lumbar and sacral
nerve roots are therefore the longest, extending far beyond the
termination of the adult cord at approximately the L2 level to reach
the remaining lumbar, sacral, and coccygeal IV foramina. This loose
bundle of spinal nerve roots arising from the lumbosacral enlargement
and the medullary cone and coursing within the lumbar cistern of CSF caudal to the termination of the spinal cord resembles a horse’s tail, hence its name—the cauda equina (L. horse tail).
is the vestigial remnant of the caudal part of the spinal cord that was
in the tail-like caudal eminence of the embryo. Its proximal end (the pial part or internal terminal filum)
consists of vestiges of neural tissue, connective tissue, and
neuroglial tissue covered by pia mater. The terminal filum perforates
the inferior end of the dural sac, gaining a layer of dura and
continuing through the sacral hiatus as the dural part or external terminal filum (also known as the coccygeal ligament)
to attach to the dorsum of the coccyx. The terminal filum is an anchor
for the inferior end of the spinal cord and the spinal meninges.
membranes that surround them are the main contents of the vertebral
canal. In adults, the spinal cord occupies only the superior two thirds
of the canal and has two (cervical and lumbosacral) enlargements
related to innervation of the limbs. The inferior, tapering end of the
cord, the medullary cone, ends at the level of the L1 or L2 vertebrae.
However, the terminal filament and the nerve roots from the lumbosacral
part of the spinal cord that form the cauda equina continue inferiorly
within the lumbar cistern containing CSF.
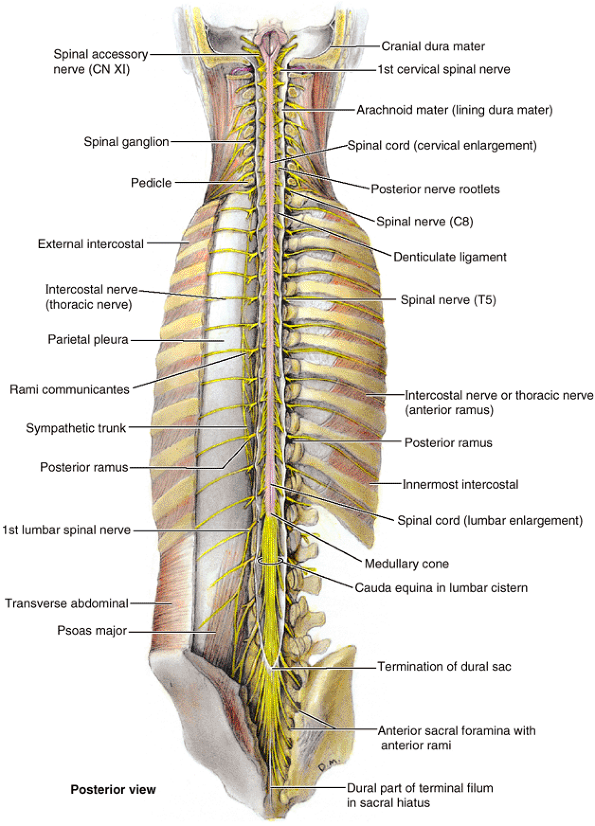 |
|
Figure 4.23. Spinal cord in situ.
The vertebral arches and the posterior aspect of the sacrum have been removed to expose the spinal cord in the vertebral canal. The spinal dural sac has also been opened to reveal the spinal cord and posterior nerve roots, the termination of the spinal cord between the L1 and the L2 vertebral level, and the termination of the spinal dural sac at the S2 segment. |
to inferior, whereas the IV foramina decrease in diameter.
Consequently, the L5 spinal nerve roots are the thickest and their
foramina, the narrowest. This increases the chance that these nerve
roots will be compressed if herniation of the nucleus pulposus occurs.
These membranes surround, support, and protect the spinal cord and
spinal nerve roots, including those of the cauda equina, and contain
the cerebrospinal fluid (CSF) in which these structures are suspended.
composed mainly of tough fibrous with some elastic tissue, is the
outermost covering membrane of the spinal cord. The spinal dura mater
is separated from the periosteum-covered bone and the ligaments that
form the walls of the vertebral canal by the epidural space. This space is occupied by the internal vertebral venous plexus embedded in a fatty matrix (epidural fat).
The epidural space runs the length of the vertebral canal, terminating
superiorly at the foramen magnum and laterally at the IV foramina, as
the dura mater adheres to the periosteum surrounding each opening, and
inferiorly, as the sacral hiatus is sealed by the sacrococcygeal
ligament.
This sac adheres to the margin of the foramen magnum of the cranium,
where it is continuous with the cranial dura mater. The spinal dural
sac is anchored inferiorly to the coccyx by the terminal filum.
The sac is evaginated by each pair of posterior and anterior roots as
they extend laterally toward their exit from the vertebral canal (Fig. 4.25). Thus tapering lateral extensions of the spinal dura surround each pair of posterior and anterior nerve roots as dural root sheaths, or sleeves (Figs. 4.24, 4.26, and 4.27). Distal to the spinal ganglia, these sheaths blend with the epineurium (outer connective tissue covering of the spinal nerves) that adheres to the periosteum lining the IV foramina.
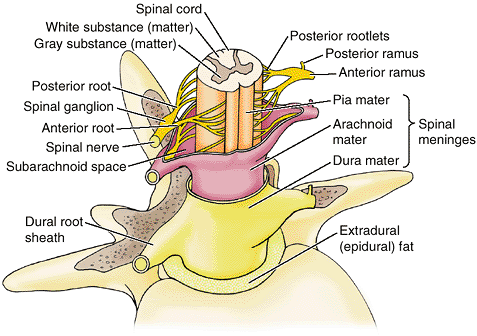 |
|
Figure 4.24. Spinal cord, spinal nerves, and spinal meninges.
Three membranes (the spinal meninges) cover the spinal cord: dura mater, arachnoid mater, and pia mater. As the spinal nerve roots extend toward an IV foramen, they are surrounded by a dural root sheath (sleeve) that is continuous distally with the epineurium of the spinal nerve. |
The function of these afferent and sympathetic fibers is unclear,
although it is known that the afferent fibers supply pain receptors
that are involved in the referred pain characteristic of spinal
disorders and become irritated when there is inflammation of the
meninges (meningitis).
is a delicate, avascular membrane composed of fibrous and elastic
tissue that lines the spinal dural sac and its dural root sheaths. It
encloses the CSF-filled subarachnoid space containing the spinal cord,
spinal nerve roots, and spinal ganglia (Figs. 4.24, 4.25, and 4.27).
The arachnoid mater is not attached to the dura mater but is held
against its inner surface by the pressure of the CSF. In a lumbar
spinal puncture, the needle traverses the dura and arachnoid mater
simultaneously. Their apposition is the dura–arachnoid interface (Table 4.4),
often erroneously referred to as the “subdural space.” No actual space
occurs naturally at this site; it is, rather, a weak cell layer (Haines, 2002). Bleeding into this layer creates a pathological space at the dura–arachnoid junction in which a subdural hematoma
is formed. In the cadaver, because of the absence of CSF, the arachnoid
mater falls away from the inner surface of the dura mater and lies
loosely on the spinal cord. The arachnoid
mater is separated from the pia mater on the surface of the spinal cord by the subarachnoid space containing CSF. Delicate strands of connective tissue, the arachnoid trabeculae, span the subarachnoid space connecting the arachnoid mater and pia mater.
|
Table 4.4. Spaces Associated with the Spinal Meninges
|
|||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||
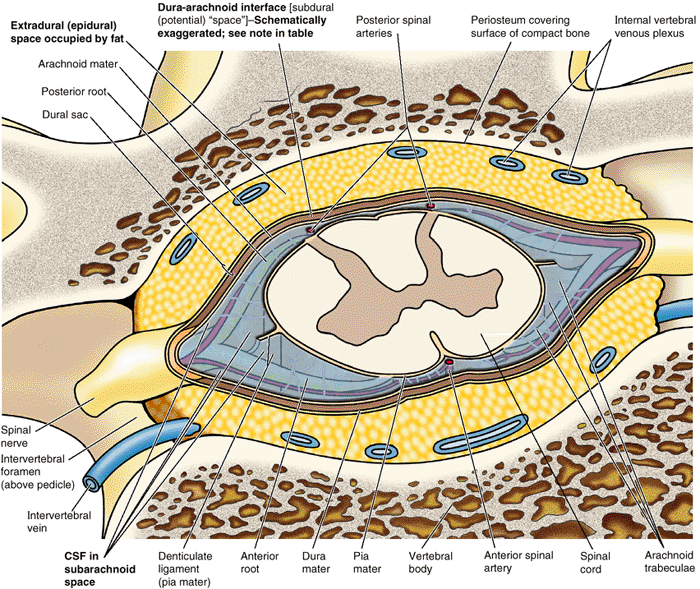
innermost covering membrane of the spinal cord, consists of flattened
cells with long, equally flattened processes that closely follow all
the surface features of the spinal cord (Haines, 2002).
The pia mater also directly covers the roots of the spinal nerves and
the spinal blood vessels. Inferior to the medullary cone, the pia mater
continues as the terminal filum (Fig. 4.22).
and 4.27).
The denticulate ligament consists of a fibrous sheet of pia mater
extending midway between the posterior and the anterior nerve roots
from the lateral surfaces of the spinal cord. The 20–22 processes,
shaped much like shark’s teeth, attach to the inner surface of the
arachnoid-lined dural sac. The superior process of each denticulate
ligament attaches to the cranial dura mater immediately superior to the
foramen magnum, and the inferior process extends from the medullary
cone, passing between the T12 and the L1 nerve roots.
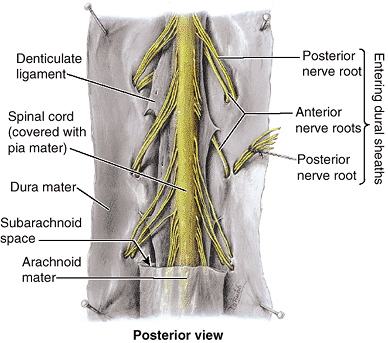 |
|
Figure 4.25. Spinal cord within its meninges. The dura mater and arachnoid mater have been split and pinned flat to expose the spinal cord and spinal nerve roots.
|
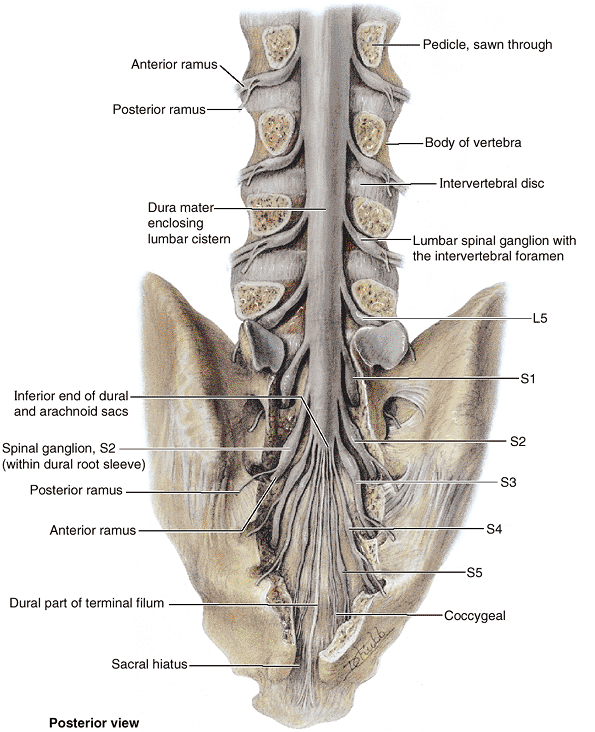 |
|
Figure 4.26. Inferior end of spinal dural sac.
A laminectomy has been performed (i.e., most of the vertebral arches of the lumbar and sacral vertebrae have been removed) to show the inferior end of the dural sac, which encloses the lumbar cistern containing CSF and the cauda equina. The lumbar spinal ganglia lie within the IV foramina, but the sacral spinal ganglia (S1–S5) are in the sacral canal. In the lumbar region, the nerves exiting the IV foramina pass superior to the IV discs at that level; thus herniation of the nucleus pulposus tends to impinge on nerves passing to lower levels. |
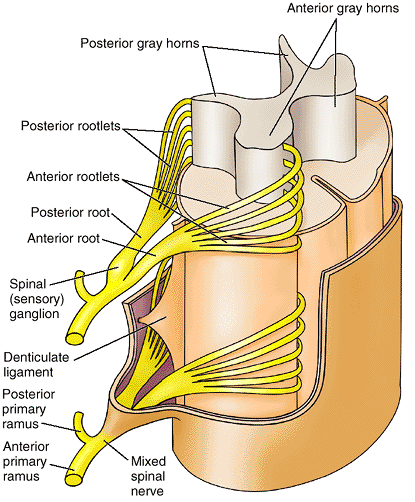 |
|
Figure 4.27. Spinal cord, anterior and posterior nerve rootlets and roots, spinal nerves, and spinal meninges.
The structure of the spinal cord (gray and white substance) and the entrance and exit of nerve fibers via rootlets are demonstrated. The denticulate ligament is a serrated shelf-like extension of the spinal pia mater projecting between the posterior and the anterior nerve roots in a frontal plane from either side of the cervical and thoracic regions of the spinal cord. Its relatively strong, tooth-like processes anchor the spinal cord within the subarachnoid space. |
The enlargement of the subarachnoid space in the dural sac, caudal to
the medullary cone and containing CSF and the cauda equina, is the lumbar cistern (Figs. 4.22 and 4.23).
It extends from the L2 vertebra to the second segment of the sacrum.
Dural root sheaths, enclosing spinal nerve roots in extensions of the
subarachnoid space, protrude from the sides of the lumbar cistern (Fig. 4.26).
(G. slender membranes). They develop as a single layer from the
mesenchyme surrounding the embryonic spinal cord. Fluid-filled spaces
form within this layer and coalesce to become the subarachnoid space (Moore and Persaud, 2003).
The origin of the pia mater and arachnoid mater from a single membrane
is reflected by the numerous arachnoid trabeculae passing between them (Table 4.4).
The arachnoid mater in adults is thick enough to be manipulated with
forceps. In contrast, the delicate pia mater is barely visible to the
unaided eye, although it gives a shiny appearance to the surface of the
spinal cord.
(spinal tap), the extraction or sampling of CSF from the lumbar
cistern, is an important diagnostic tool for evaluating a variety of
central nervous system (CNS) disorders. Meningitis
and diseases of the CNS may alter the cells in the CSF or change the
concentration of its chemical constituents. Examination of CSF can also
determine if blood is present. Lumbar spinal puncture is performed with
the patient leaning forward or lying on the side with the back flexed (Fig. B4.17).
Flexion of the vertebral column facilitates insertion of the needle by
spreading the vertebral laminae and spinous processes apart, stretching
the ligamenta flava.
fitted with a stylet, is inserted in the midline between the spinous
processes of the L3 and L4 (or L4 and L5) vertebrae. Recall that a
plane transecting the highest points of the iliac crests—the supracristal plane—usually
passes through the L4 spinous process. At these levels, there is no
danger of damaging the spinal cord. After passing 4–6 cm in adults
(more in obese persons), the needle punctures the dura mater and
arachnoid mater and enters the lumbar cistern. When the stylet is
removed, CSF escapes at the rate of approximately one drop per second.
If subarachnoid pressure is high, CSF flows out or escapes as a jet.
Lumbar puncture is not performed in the presence of increased
intracranial pressure. Intracranial pressure is now generally
determined through CT scanning before lumbar puncture, but it may also
be determined by examination of the fundus (back) of the interior of
the eyeball with an ophthalmoscope (see clinical correlation [blue] box “Papilledema,” in Chapter 7).
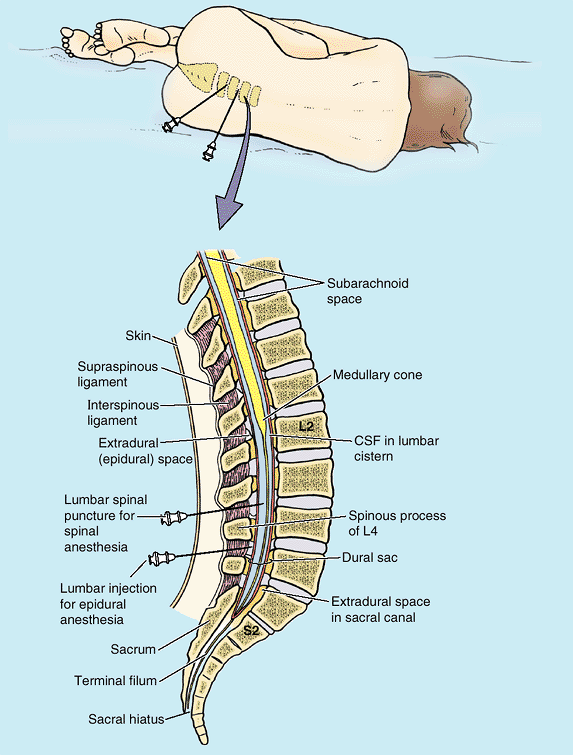 |
|
Figure B4.17
|
which likely results from the leakage of CSF through the lumbar
puncture. The clinical anatomy of spinal blocks for childbirth is
discussed in Chapter 3.
(epidural) space using the position described in clinical correlation
(blue) box “Lumbar Spinal Puncture,” (lumbar epidural block), or
through the sacral hiatus (caudal epidural block). Epidural block for
anesthesia during childbirth is discussed in Chapter 3.
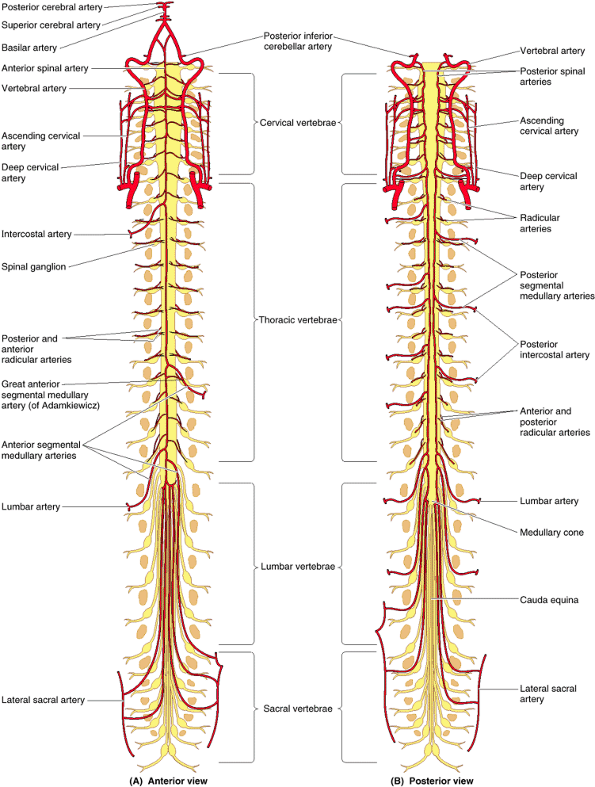 |
|
Figure 4.28. Arterial supply of spinal cord. A and B.
The arteries derive from branches of the vertebral, ascending and deep cervical, intercostal, lumbar, and lateral sacral arteries. Three longitudinal arteries supply the spinal cord: an anterior spinal artery and two posterior spinal arteries. These vessels are reinforced by blood from the anterior and posterior segmental medullary arteries. Segmental medullary arteries supply the nerve roots that they course along and then contribute to the longitudinal arteries. The great anterior segmental medullary artery (of Adamkiewicz) is the largest segmental medullary artery, vital to the blood supply of the spinal cord between its cervical and lumbosacral enlargements. At levels where there are no segmental medullary arteries, radicular arteries supply the posterior and anterior roots of the spinal nerves. (Radicular arteries are shown at only the cervical and thoracic levels, but they also occur at the lumbar and sacral levels.) |
vertebral canal are suspended in CSF contained within the dural sac and
sheaths. The fluid-filled subarachnoid space is lined with pia mater
and arachnoid mater, which are continuous membranes. Because the spinal
cord does not extend into the lumbar cistern (the inferior part of the
subarachnoid space), it is an ideal site for sampling CSF or for
injection of anesthetic agents.
the vertebral, ascending cervical, deep cervical, intercostal, lumbar,
and lateral sacral arteries (Figs. 4.28 & 4.29). Three longitudinal arteries supply the spinal cord: an anterior spinal artery and paired posterior spinal arteries. These arteries run longitudinally from the medulla of the brainstem to the medullary cone of the spinal cord. The anterior spinal artery, formed by the union of branches of the vertebral arteries, runs inferiorly in the anterior median fissure. Sulcal arteries arise from the anterior spinal artery and enter the spinal cord through this fissure (Fig. 4.29B). The sulcal arteries supply approximately two thirds of the cross-sectional area of the spinal cord (Williams et al., 1995). Each posterior spinal artery is a branch of either the vertebral artery or the posteroinferior cerebellar artery (Figs. 4.28B and 4.29). The posterior spinal arteries commonly form anastomosing channels in the pia mater.
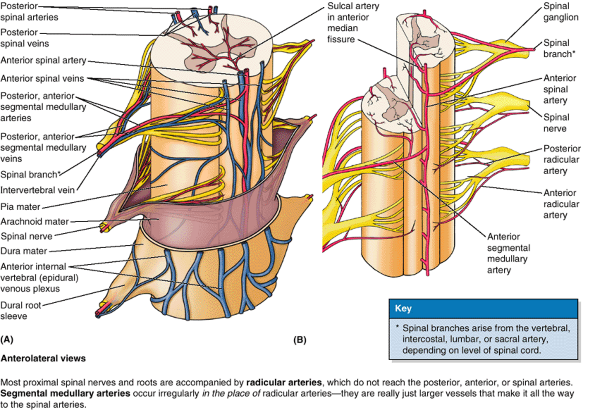 |
|
Figure 4.29. Arterial supply and venous drainage of spinal cord and spinal nerve roots. A.
The veins that drain the spinal cord, as well as the internal vertebral venous plexuses, drain into the intervertebral veins, which in turn drain into segmental veins. B. The pattern of the arterial supply of the spinal cord is from three longitudinal arteries: one anterior lying in the anteromedian position and the other two lying posterolaterally. These vessels are reinforced by medullary branches derived from the segmental arteries. The sulcal arteries are small branches of the anterior spinal artery coursing in the anterior median fissure. |
arteries can supply only the short superior part of the spinal cord.
The circulation to much of the spinal cord depends on segmental
medullary and radicular arteries running along the spinal nerve roots.
The anterior and posterior segmental
medullary arteries
are derived from spinal branches of the ascending cervical, deep
cervical, vertebral, posterior intercostal, and lumbar arteries. The
segmental medullary arteries are located chiefly where the need for a
good blood supply to the spinal cord is greatest—the cervical and
lumbosacral enlargements. They enter the vertebral canal through the IV
foramina.
(of Adamkiewicz), which is on the left side in about 65% of people,
reinforces the circulation to two thirds of the spinal cord, including
the lumbosacral enlargement (Fig. 4.28A).
The great anterior segmental medullary artery, much larger than the
other segmental medullary arteries, usually arises from an inferior
intercostal or upper lumbar artery and enters the vertebral canal
through the IV foramen at the lower thoracic or upper lumbar level.
Most spinal nerve roots and proximal spinal nerves and roots are
accompanied by radicular arteries that do not reach the posterior,
anterior, or spinal arteries. Segmental medullary arteries occur
irregularly in the place of radicular arteries; they are really larger
vessels that make it all the way to the spinal arteries. Most radicular
arteries are small and supply only the nerve roots; however, some of
them assist with the supply of superficial parts of the gray matter in
the posterior and anterior horns of the spinal cord.
distribution similar to that of the spinal arteries. There are usually
three anterior and three posterior spinal veins (Fig. 4.29A). The spinal veins are arranged longitudinally, communicate freely with each other, and are drained by up to 12 anterior and posterior medullary and radicular veins. The veins draining the spinal cord join the internal vertebral (epidural) venous plexus in the epidural space (Fig. 4.20).
The internal vertebral venous plexus passes superiorly through the
foramen magnum to communicate with dural sinuses and vertebral veins in
the cranium (see Chapter 7). The internal
vertebral plexus also communicates with the external vertebral venous
plexus on the external surface of the vertebrae.
are reinforced by irregular and asymmetric segmental medullary arteries
that also supply the spinal nerve roots they course along. Radicular
arteries supply the remainder of the nerve roots. The veins draining
the spinal cord have a distribution and drainage generally reflective
of the spinal arteries, although there are normally three longitudinal
spinal veins both anteriorly and posteriorly.
spinal cord from the segmental medullary arteries are important in
supplying blood to the anterior and posterior spinal arteries.
Fractures, dislocations, and fracture dislocations may interfere with
the blood supply to the spinal cord from the spinal and medullary
arteries.
cord affects its function and can lead to muscle weakness and
paralysis. The spinal cord may also suffer circulatory impairment if
the segmental medullary arteries, particularly the great anterior
segmental medullary artery (of Adamkiewicz), are narrowed by obstructive arterial disease.
Sometimes the aorta is purposely occluded (cross clamped) during
surgery. Patients undergoing such surgeries, and those with ruptured
aneurysms of the aorta or occlusion of the great anterior segmental
medullary artery, may lose all sensation and voluntary movement
inferior to the level of impaired blood supply to the spinal cord (paraplegia) secondary to death of neurons in the part of the spinal cord supplied by the anterior spinal artery (Figs. 4.28 and 4.29).
Neurons with cell bodies distant from the site of ischemia will also be
lost secondary to the degeneration of axons traversing the site. The
likelihood of iatrogenic paraplegia
depends on such factors as the age of the patient, the extent of the
disease, and the length of time the aorta is cross clamped (Murray et al., 1992).
blood flow from the segmental medullary arteries to the anterior spinal
artery supplying the midthoracic region of the spinal cord may be
reduced or stopped. These people may also lose sensation and voluntary
movement in the areas supplied by the affected level of the spinal cord.
shape from level to level, particularly in the cervical and lumbar
regions. A narrow vertebral canal in the cervical region, into which
the spinal cord fits tightly, is potentially dangerous because a minor
fracture and/or dislocation of a cervical vertebra may damage the
spinal cord. The protrusion of a cervical IV disc into the vertebral
canal after a neck injury may cause spinal cord shock
associated with paralysis inferior to the site of the lesion. In some
people, no fracture or dislocation of cervical vertebrae can be found.
If the individual dies and an autopsy is performed, a softening of the
spinal cord may be detected at the site of the cervical disc
protrusion. Encroachment of the vertebral canal by a protruding IV
disc, by swollen ligamenta flava, or resulting from osteoarthritis of the zygapophysial joints
may exert pressure on one or more of the spinal nerve roots of the
cauda equina. Pressure may produce sensory and motor symptoms in the
area of distribution of the involved spinal nerve. This group of bone
and joint abnormalities, called lumbar spondylosis (degenerative joint disease), also causes localized pain and stiffness.
results in loss of all sensation and voluntary movement inferior to the
lesion. Transection of the spinal cord between the following levels
will result in the indicated effects:
-
C1–C3: The patient has no function below head level; a ventilator is required to maintain respiration (otherwise fatal).
-
C4–C5: Respiration occurs, but the patient is quadriplegic (no function of upper or lower limbs).
-
C6–C8: The patient experiences a loss of
hand and a variable amount of upper limb function; the individual may
be able to self-feed or propel a wheelchair. -
T1–T9 (between cervical and lumbosacral
enlargements): The individual is paraplegic (paralysis of both lower
limbs); the amount of trunk control varies with the height of the
lesion. -
T10–L1: The patient has some thigh muscle function, which may allow walking with long leg braces.
-
L2–L3: The individual retains most leg muscle function; short leg braces may be required for walking.
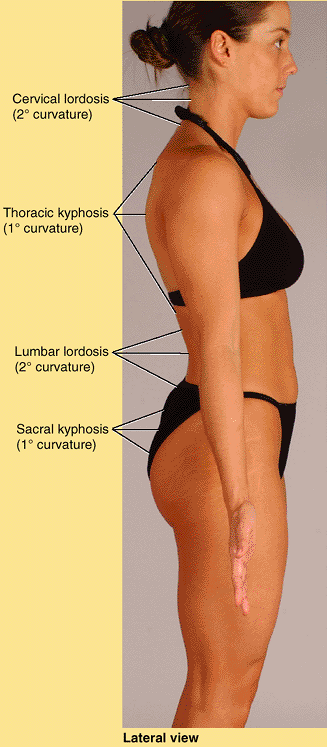 |
|
Figure SA4.1. Normal curvatures of back.
|
is the most superior cervical spinous process that is usually evident
superficially. Often, when the patient stands erect, this is the only
spinous process visible; hence the name vertebra prominens. The spinous process of C2 is the first bony point that can be felt deeply in the midline inferior to the external occipital protuberance,
a median projection located a little inferior to the bulging part of
the posterior aspect of the head at the junction of the head and neck.
C1 has no spinous process, and its small posterior tubercle is neither
visible nor palpable. The short bifid spinous processes of the C3–C5
vertebrae may be felt in the nuchal groove
between the neck muscles, but they are not easy to palpate because the
cervical curvature, which is concave posteriorly, places them deep to
the surface from which they are separated by the nuchal ligament. The
bifid spinous process of C6 vertebra is
easily felt superior to the visible tip of the C7 process when the neck
is flexed because of its long process. When the neck and back are
flexed, the spinous processes of the upper thoracic vertebra may also be seen. If the individual is especially lean, a continuous ridge appears linking their tips—the supraspinous ligament.
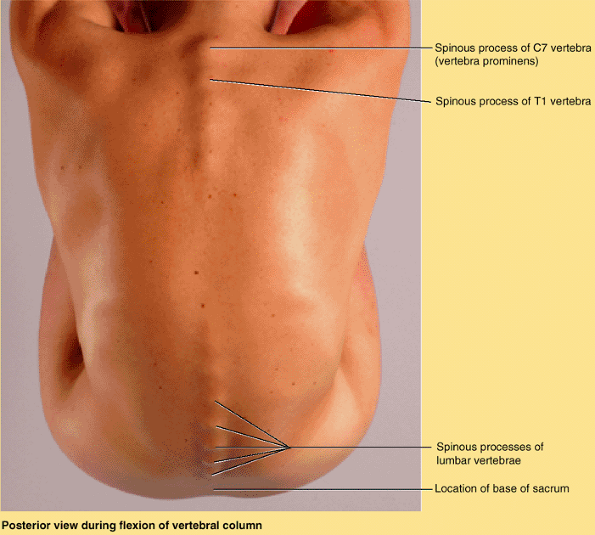 |
|
Figure SA4.2. Surface anatomy of back during flexion.
|
and readily palpable, the spinous process of T1 may actually be the
most prominent. The spinous processes of the other thoracic vertebrae
may be obvious in thin people and in others can be identified by
superior to inferior palpation beginning at the C7 spinous process,
immediately superior to the T1 spinous process. The tips of the thoracic spinous processes do not indicate the level of the corresponding vertebral bodies because they overlap (lie at the level of) the vertebra below (Fig. 4.1D).
The tips of the spinous processes are normally in line with each other,
even if the collective line wanders slightly from the midline. A sudden
shift in the alignment of adjacent spinous processes may be the result
of a unilateral dislocation of a zygapophysial joint; however, slight
irregular misalignments may also result from a fracture of the spinous
process. The short 12th rib, the lateral end of which can be palpated
in the posterior axillary line, can be used to confirm identity of the
T12 spinous process.
lies anterior to it. In most people, the transverse processes of
thoracic vertebrae can be palpated on each side of the spinous
processes in the thoracic region; and in lean individuals, the ribs can
be palpated from tubercle to angle, at least in the lower back
(inferior to the scapula).
provides an estimate of the position of the inferior end of the spinal
cord. A horizontal line joining the highest points of the iliac crests
passes through the tip of the L4 spinous process and the L4–L5 IV disc (Fig. SA4.3B). This is a useful landmark when performing a lumbar puncture to obtain a sample of CSF (see clinical correlation [blue] box “Lumbar Spinal Puncture,” in this chapter).
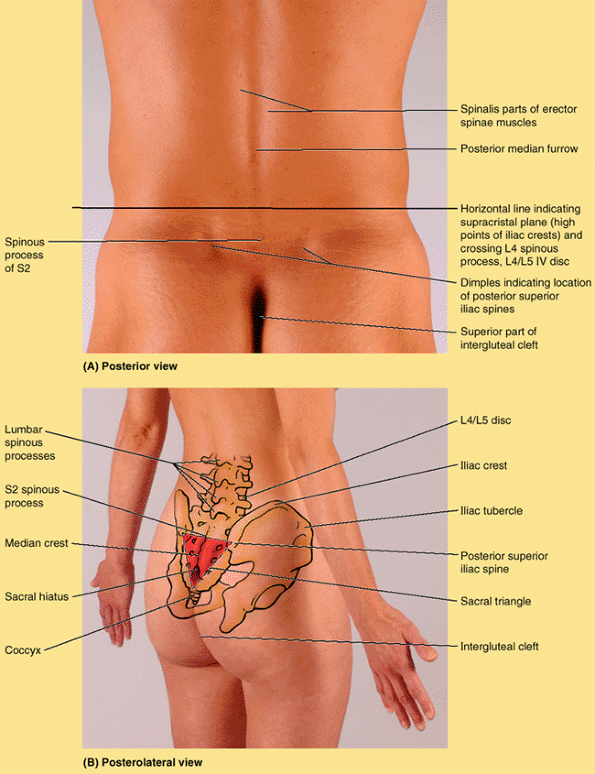 |
|
Figure SA4.3. Surface anatomy of lower back.
|
formed by the fusion of the spinous processes of the superior three or
four sacral vertebrae, can be felt inferior to the L5 spinous process (Fig. SA4.3B). The sacral triangle is formed by the lines joining the two posterior superior iliac spines and the superior part of the intergluteal (natal) cleft between the buttocks. The sacral triangle outlining the sacrum is a common area of pain resulting from low back sprains. The sacral hiatus can be palpated at the inferior end of the sacrum located in the superior part of the intergluteal cleft (Fig. B4.6).
can be palpated approximately 2.5 cm posterosuperior to the anus.
Clinically, the coccyx is examined with a gloved finger in the anal
canal.
especially in obese people; consequently, the many strong muscles
attached to the spinous and transverse processes are necessary to
support and move the vertebral column. There are two major groups of
muscles in the back. The extrinsic back muscles include superficial and intermediate muscles that produce and control limb and respiratory movements, respectively. The intrinsic (deep) back muscles include muscles that specifically act on the vertebral column, producing its movements and maintaining posture.
(trapezius, latissimus dorsi, levator scapulae, and rhomboids) connect
the upper limbs to the trunk and produce and control limb movements (Fig. 4.30A; see also Chapter 6).
Although located in the back region, for the most part these muscles
receive their nerve supply from the anterior rami of cervical nerves
and act on the upper limb. The trapezius receives its motor fibers from
a cranial nerve, the spinal accessory nerve (CN XI).
(serratus posterior) are thin muscles, commonly designated superficial
respiratory muscles, but are more likely proprioceptive rather than
motor in function (Vilensky et al., 2001). They are described with muscles of the thoracic wall (see Chapter 1). The serratus posterior superior lies deep to the rhomboids, and the serratus posterior inferior
lies deep to the latissimus dorsi. Both serratus muscles are innervated
by intercostal nerves, the superior by the first four intercostals and
the inferior by the last four.
deep back muscles) are innervated by the posterior rami of spinal
nerves and act to maintain posture and control movements of the
vertebral column (Figs. 4.30B and 4.31).
These muscles, which extend from the pelvis to the cranium, are
enclosed by deep fascia that attaches medially to the nuchal ligament,
the tips of the spinous processes of the vertebrae, the supraspinous
ligament, and the median crest of the sacrum. The fascia attaches
laterally to the cervical and lumbar transverse processes and to the
angles of the ribs. The thoracic and lumbar parts of the deep fascia
constitute the thoracolumbar fascia. It
extends laterally from the spinous processes and forms a thin covering
for the deep muscles in the thoracic region and a strong thick covering
for muscles in the lumbar region. The deep back muscles are grouped
into superficial, intermediate, and deep layers according to their
relationship to the surface (Tables 4.5, 4.6 and 4.7).
are thick and flat and lie on the lateral and posterior aspects of the
neck, covering the vertical muscles somewhat like a bandage, which
explains their name (L. splenion, bandage) (Fig. 4.31). The splenii arise from the midline and extend superolaterally to the cervical vertebrae (splenius cervicis) and cranium (splenius capitis).
The splenii cover and hold the deep neck muscles in position.
Information on the attachments, nerve supply, and actions of the
superficial layer of intrinsic muscles is provided in Table 4.5.
lie in a “groove” on each side of the vertebral column between the
spinous processes centrally and the angles of the ribs laterally (Fig. 4.31). The massive erector spinae is the chief extensor of the vertebral column and is divided into three columns: The iliocostalis forms the lateral column, the longissimus forms the intermediate column, and the spinalis
forms the medial column. Each column is divided regionally into three
parts according to the superior attachments (e.g., iliocostalis
lumborum, iliocostalis thoracis, and iliocostalis cervicis). The common
origin of the three erector spinae columns is through a broad tendon
that attaches inferiorly to the posterior part of the iliac crest, the
posterior aspect of the sacrum, the sacroiliac ligaments, and
the
sacral and inferior lumbar spinous processes. The erector spinae are
often referred to as the “long muscles” of the back. In general, they
are dynamic (motion-producing) muscles, acting bilaterally to extend
(straighten) the flexed trunk. Information on the attachments, nerve
supply, and actions of the intermediate layer of intrinsic muscles is
provided in Table 4.6.
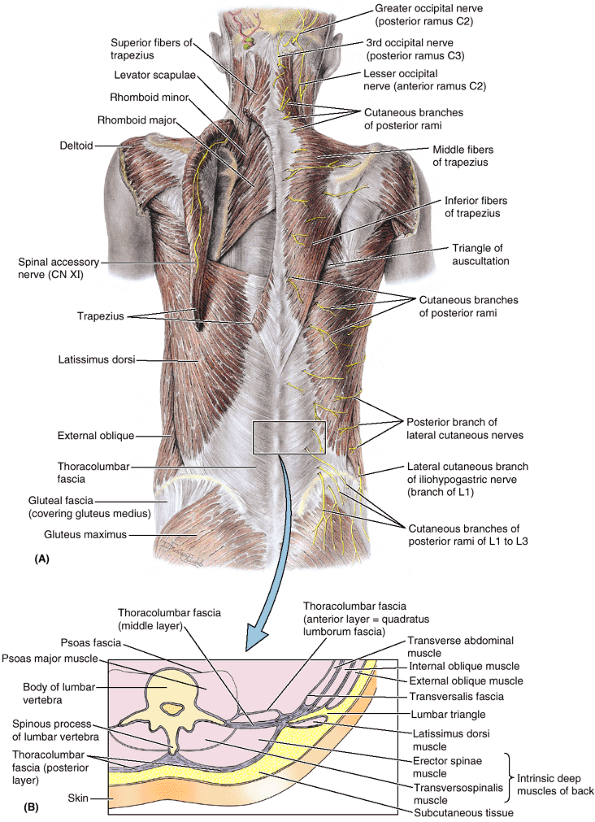 |
|
Figure 4.30. Muscles of back. A.
The superficial extrinsic muscles are shown. The trapezius is reflected on the left to show the spinal accessory nerve (CN XI), coursing on its deep surface, and the levator scapulae and rhomboid muscles. These muscles help attach the upper limb to the trunk. B. This transverse section of part of the back shows the location of the intrinsic back muscles and the layers of fascia associated with them. The combined posterior aponeuroses of the transverse abdominal and internal oblique muscles splits into two strong sheets, the middle and posterior layers of the thoracolumbar fascia, which enclose the intrinsic back muscles. |
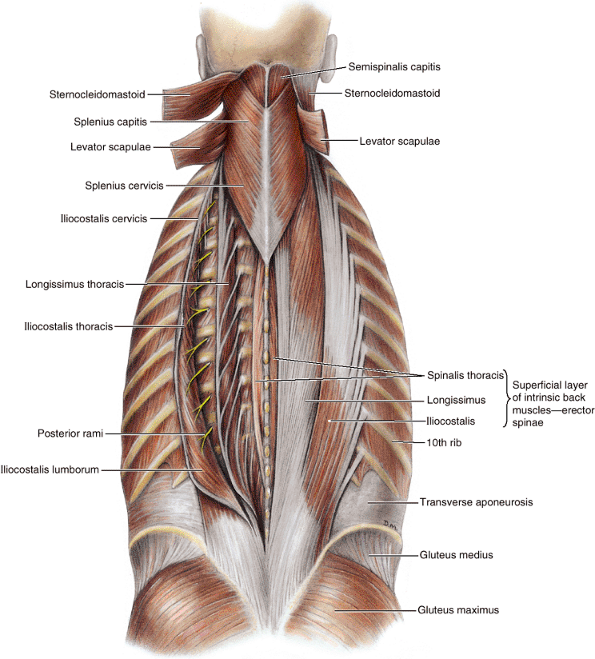 |
|
Figure 4.31. Superficial and intermediate layers of intrinsic back muscles: splenius and erector spinae.
The sternocleidomastoid and levator scapulae muscles are reflected to reveal the splenius capitis and splenius cervicis muscles. On the right side, the erector spinae is undisturbed (in situ) and shows the three columns of this massive muscle. On the left side, the spinalis muscle, the thinnest and most medial of the erector spinae columns, is displayed as a separate muscle by reflecting the longissimus and iliocostalis columns of the erector spinae. |
|
Table 4.5. Superficial Layer of Intrinsic Back Muscles
|
|||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||
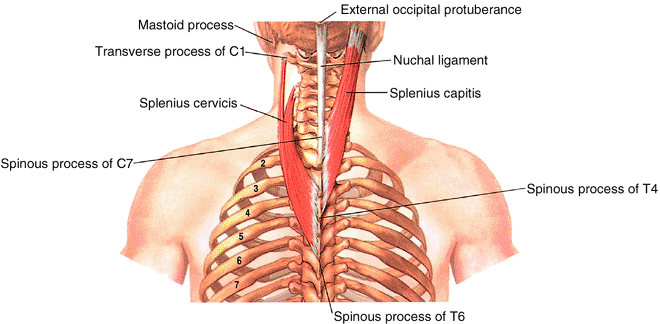
consisting of the semispinalis, multifidus, and rotatores. These
muscles originate from transverse processes of vertebrae and pass to
spinous processes of more superior vertebrae. They occupy the “gutter”
between the transverse and the spinous processes and are attached to
these processes, the laminae between them, and the ligaments linking
them together (Fig. 4.32).
superficial member of the group. As its name indicates, it arises from
approximately half of the vertebral column (spine) (Fig. 4.31). It is divided into three parts according to the superior attachments: semispinalis capitis, semispinalis thoracis, and semispinalis cervicis. Semispinalis capitis is responsible for the longitudinal bulge in the back of the neck near the median plane. The multifidus is the middle layer of the group and consists of short, triangular muscular bundles that are thickest in the lumbar region (Fig. 4.32A). The rotatores,
or rotator muscles, are the deepest of the three layers of
transversospinal muscles and are best developed in the thoracic region.
Details concerning the attachments, innervation, and action of the
transversospinalis group of the deep layer of intrinsic back muscles
are provided in Table 4.7.
are minor deep back muscles that are poorly developed in the thoracic
region. The interspinal and intertransverse muscles connect spinous and
transverse processes, respectively. The elevators of the ribs represent
the posterior intertransverse muscles of the neck. Details concerning
the attachments, nerve supply, and actions of the minor muscles of the
deep layer of intrinsic muscles are provided in Table 4.7.
The back muscles are relatively inactive in the stand-easy position,
but they (especially the shorter deep layer of intrinsic muscles) act
as static postural muscles (fixators, or steadiers) of the vertebral
column, maintaining tension and stability as required for the erect
posture. Note in Table 4.9 that all movements
of the IV joints (i.e., all movements of the vertebral column) except
pure extension involve or are solely produced by the concentric contraction of abdominal muscles. However, bear in mind that in these as in all movements,
the eccentric contraction (controlled relaxation) of the antagonist muscles is vital to smooth, controlled movement (see “Muscle Tissue and the Muscular System” in the Introduction).
Thus it is actually the interaction of anterior (abdominal) and
posterior (back) muscles (as well as the contralateral pairs of each)
that provides the stability and produces motion of the axial skeleton,
much like guy (guide) wires support a pole. Often chronic back strain
(such as that caused by excessive lumbar lordosis) results from
imbalance in this support (lack of tonus of abdominal muscles in the
case of lordosis). To restore that balance, the patient may need to
exercise or eliminate excessive, unevenly distributed weight.
|
Table 4.6. Intermediate Layer of Intrinsic Back Muscles
|
|||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||
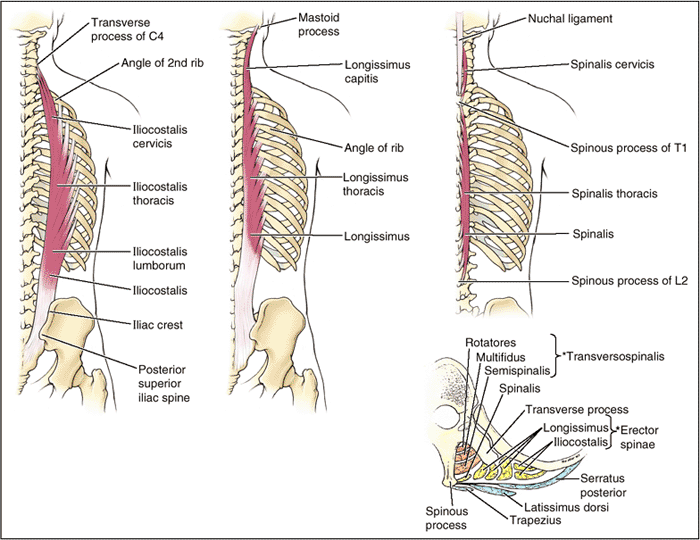
that the higher concentration of spindles occurred because small
muscles produce the most precise movements, such as fine postural
movements or manipulation and, therefore, require more proprioceptive
feedback. The movements described for small muscles are deduced from
the location of their attachments and the direction of the muscle
fibers and from activity measured by electromyography as movements are
performed. Muscles such as the rotatores, however, are so small and are
placed in positions of such relatively poor mechanical advantage that
their ability to produce the movements described is somewhat
questionable. Furthermore, such small muscles are often redundant to
other larger muscles that have superior mechanical advantage. Hence it
has been proposed (Buxton and Peck, 1989)
that the smaller muscles of small–large muscle pairs function more as
“kinesiological monitors,” or organs of proprioception and that the
larger muscles are the producers of motion.
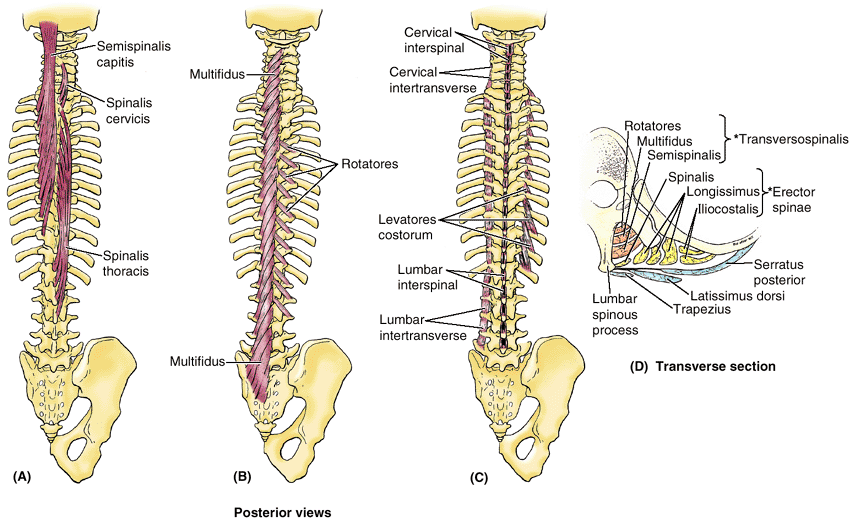
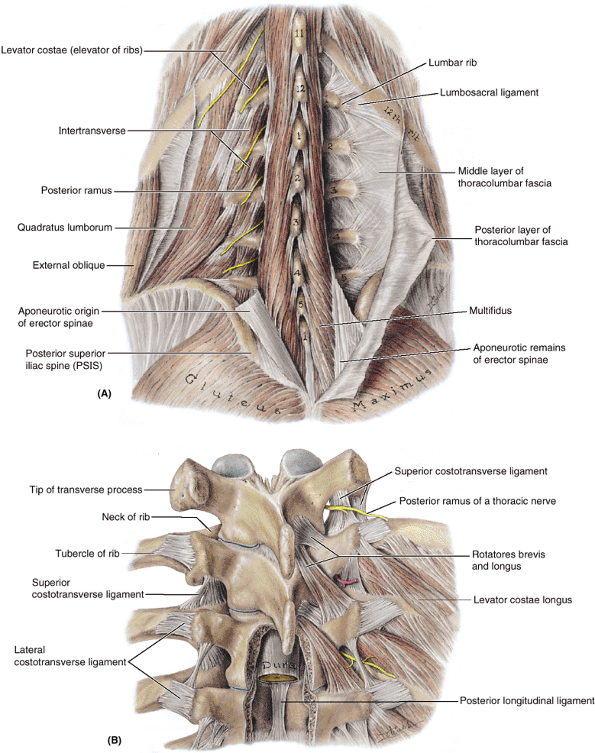 |
|
Figure 4.32. Muscles of deep layer of intrinsic back muscles. A.
The transversospinal muscle group is deep to the erector spinae. It consists chiefly of a large number of small muscles that run obliquely upward and medially from transverse to spinous processes. The multifidus and quadratus lumborum muscles are shown as is the thoracolumbar fascia. The short lumbar rib is articulating with the transverse process of L1 vertebra. This common variation does not usually cause a problem; however, those unfamiliar with its possible presence may think it is a fractured transverse process. B. Deeper dissection showing the rotatores and costotransverse ligaments. Two of the three sets of costotransverse ligaments are seen: superior and lateral. The (medial) costotransverse ligaments extend between the neck of the rib and the transverse process of the vertebra of the same number (not shown). The levator muscles, which elevate the ribs, represent the posterior intertransverse muscles in the thoracic region. |
|
Table 4.8. Principal Muscles Producing Movement of the Cervical Intervertebral Joints
|
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||

|
Table 4.9. Principal Muscles Producing Movements of the Thoracic and Lumbar Intervertebral (IV) Joints
|
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||
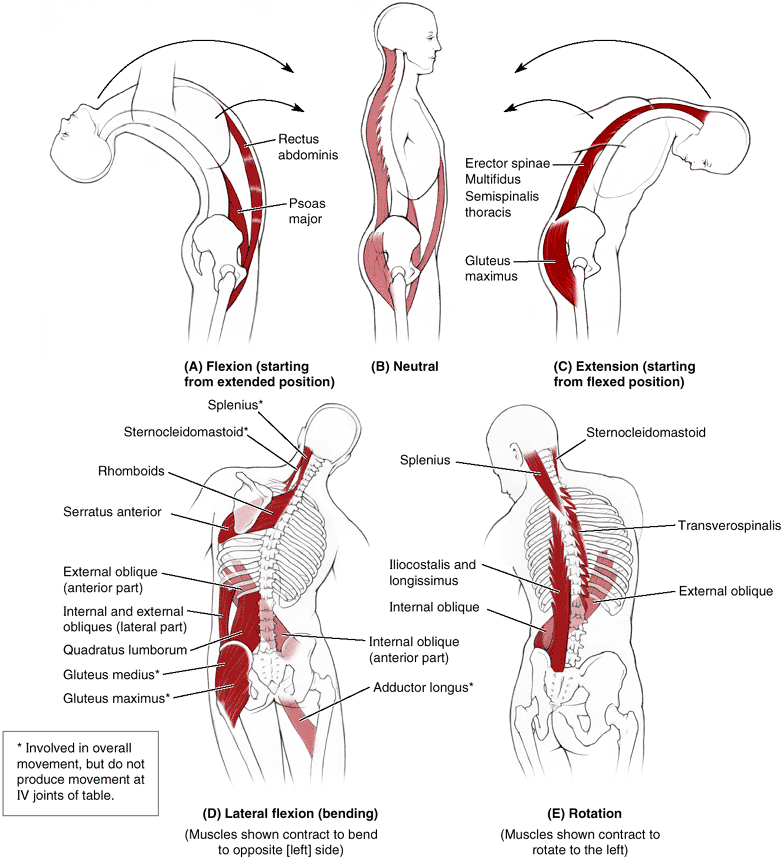
Deep (intrinsic) back muscles provide primarily extension and
proprioception for posture and work synergistically with the muscles of
the anterolateral abdominal wall to produce movements of the trunk.
is an injury in which only ligamentous tissue, or the attachment of
ligament to bone, is involved, without dislocation or fracture. It
results from excessively strong contractions related to movements of
the vertebral column, such as excessive extension or rotation. Back strain
is a common back problem in people who participate in sports; it
results from overly strong muscular contraction. Back strain involves
some degree of stretching or microscopic tearing of muscle fibers. The
muscles usually involved are those producing movements of the lumbar IV
joints, especially the erector spinae columns. If the weight is not
properly balanced on the vertebral column, strain is exerted on the
muscles. This is undoubtedly a common cause of low back pain.
strain on the vertebral column and its ligaments and muscles. These
strains can be minimized if the lifter crouches, holds the back as
straight as possible, and uses the muscles of the buttocks and lower
limbs to assist with the lifting. As a protective mechanism, the back
muscles go into spasm after an injury or
in response to inflammation of structures in the back, such as
ligaments. A spasm is a sudden involuntary contraction of one or more
muscle groups. Spasms are attended by cramps, pain, and interference
with function, producing involuntary movement and distortion.
produce prominent vertical bulges on each side of the furrow. In the
lumbar region, the erector spinae are readily palpable, and their
lateral borders are indicated by shallow grooves in the skin. When the
individual is standing, the lumbar spinous processes may be indicated
by depressions in the skin. These processes usually become visible when
the vertebral column is flexed (Fig. SA4.2). The posterior median furrow ends in the flattened triangular area covering the sacrum and is replaced inferiorly by the intergluteal cleft.
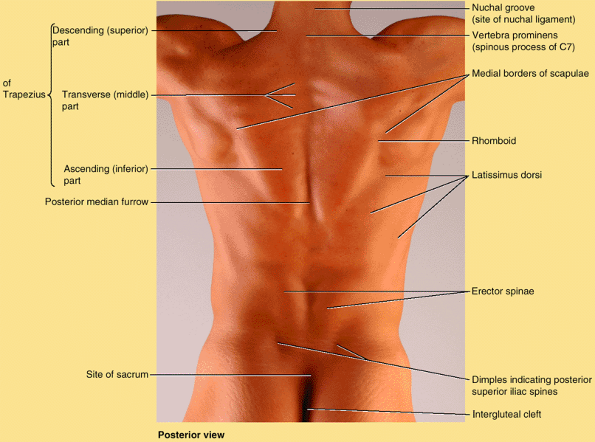 |
|
Figure SA4.4. Surface anatomy of muscles of back.
|
superior part of the posterior cervical region, is the triangular area
inferior to the occipital region of the head, including the posterior
aspects of vertebrae C1 and C2. The four small muscles of the
suboccipital region lie deep to the trapezius and semispinalis capitis
muscles and consist of two rectus capitis posterior (major and minor)
and two obliquus muscles. All four muscles are innervated by the posterior ramus of C1, the suboccipital nerve. The nerve emerges as the vertebra artery courses deeply between the occipital bone and the atlas (vertebra C1) within the suboccipital triangle.
Details concerning the boundaries and contents of this triangle and the
attachments of the suboccipital muscles are provided and illustrated in
Table 4.10.
is the only “capitis” muscle that has no attachment to the cranium.
These muscles are mainly postural muscles, but actions are typically
described for each muscle in terms of producing movement of the head.
The muscles are considered to act on the head directly or indirectly
(explaining the inclusion of capitis in
their names) by extending it on vertebra C1 and rotating it on
vertebrae C1 and C2. However, recall the discussion of the small member
of the small–large muscle pair functioning as a kinesiological monitor
for the sense of proprioception. The principal muscles producing
movements of the craniovertebral joints are summarized in Tables 4.11 and 4.12, and the nerves of the suboccipital region and posterior neck are summarized in Table 4.13.
|
Table 4.10. Suboccipital Muscles and Suboccipital Triangle
|
|||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||||||||||||||||||||||||||||||||
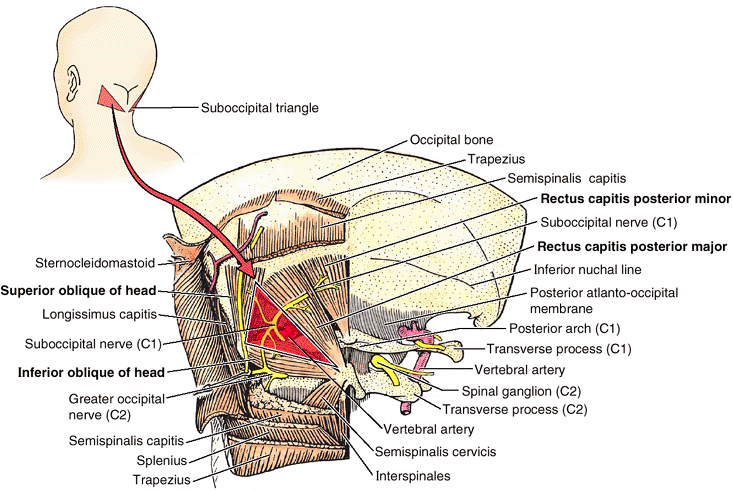
|
Table 4.11. Principal Muscles Producing Movement of the Atlanto-Occipital Joints
|
|||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||||||||
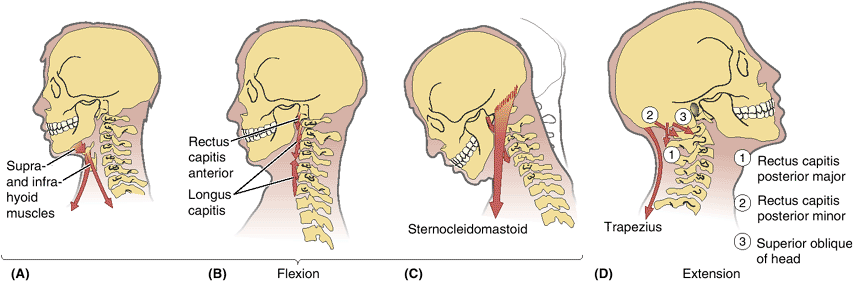
|
Table 4.12. Principal Muscles Producing Movement of the Atlanto-Axial Jointsa
|
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||
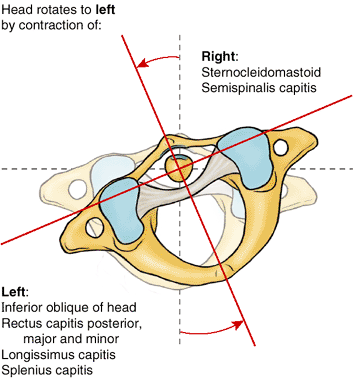
|
Table 4.13. Nerves of the Suboccipital Region/Posterior Neck
|
||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||||||||||||||
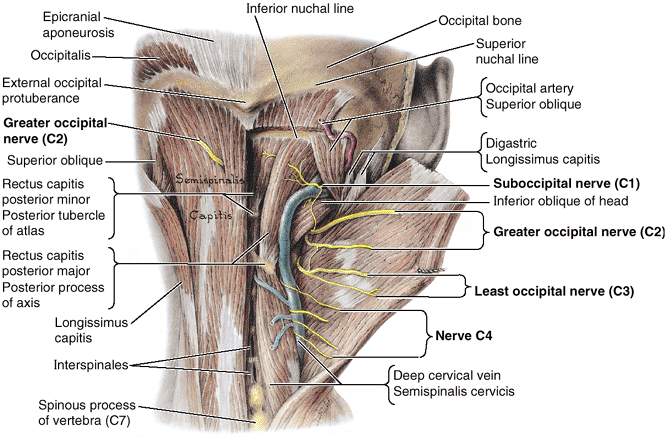
transverse foramina of the transverse processes of the cervical
vertebrae and through the suboccipital triangles becomes clinically
significant when blood flow through these arteries is reduced, as
occurs with arteriosclerosis. Under these
conditions, prolonged turning of the head, as occurs when backing up a
motor vehicle, may cause light-headedness, dizziness, and other
symptoms from the interference with the blood supply to the brainstem.
usually requires both AP and lateral views. The anatomical features
visible in the regions of the vertebral column are illustrated and
described in Figures 4.33, 4.34, 4.35 and 4.36. Conventional radiographs are excellent for high contrast structures such as bone. The advent of digital radiography allows improved contrast resolution.
is a common metabolic bone disease that is often detected during
routine radiographical studies. Vertical striping may become apparent,
reflecting the loss of the horizontal supporting trabeculae and
thickening of the vertical struts (Fig. 4.37A).
Radiographs taken during early to moderate osteoporosis demonstrate
demineralization, which is evident as diminished radiodensity of the
trabecular (spongy) bone of the vertebral bodies, causing the thinned
cortical bone to appear relatively prominent (Fig. 4.37B). Radiographs in later stages may reveal vertebral collapse (compression fractures) and increased thoracic kyphosis (Fig. 4.37C).
Vertebral body osteoporosis occurs in all vertebrae but is most common
in thoracic vertebrae and is an especially common finding in
postmenopausal females.
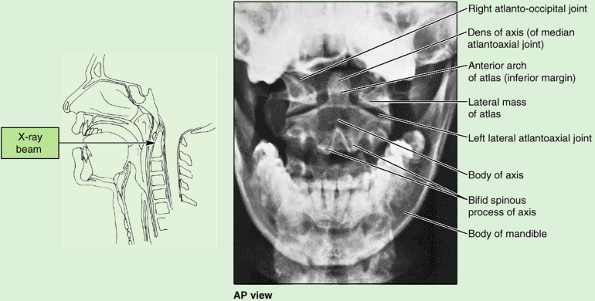 |
|
Figure 4.33. Radiograph of superior part of cervical region of vertebral column.
As shown in the orientation drawing, the radiograph was taken through the open mouth. The lateral atlantoaxial joint and the body of the axis (C2 vertebra), with its dens projecting superiorly between the lateral masses of the atlas (C1 vertebra), are seen. |
In this procedure, CSF is withdrawn by lumbar puncture and replaced
with a contrast material injected into the spinal subarachnoid space.
This technique shows the extent of the subarachnoid space and its
extensions around the spinal nerve roots within the dural root sheaths.
High-resolution MRI has largely supplanted myelography (McCormick et al., 2000).
the brain and spinal cord. It has also improved the radiological
assessment of fractures of the vertebral column, particularly in
determining the degree of compression of the spinal cord (McCormick, 2000).
CT images of the vertebral column are used to detect fractures,
lesions, and congenital anomalies. The dense vertebrae attenuate much
of the X-ray beam and, therefore, appear white on the scans (Fig. 4.39).
The IV discs have a higher density than the surrounding adipose tissue
in the epidural space and the CSF in the subarachnoid space.
Herniations of the IV discs are thus recognizable in CT images, as are
displaced fragments of the IV discs.
but X-rays are not used, as with CT. MRI produces extremely good images
of the vertebral column, spinal cord, and CSF (Fig. 4.40).
Until relatively recently, a disadvantage of MRI was that the patient
had to remain motionless in the scanner for long periods; however, new
technologies have markedly decreased this time. MRI clearly
demonstrates the components of the IV discs and shows their
relationship to the vertebral bodies and longitudinal ligaments.
Herniations of the nucleus pulposus and its relationship to the spinal
nerve roots are also well defined (Fig. 4.41).
MRI is the imaging procedure of choice for evaluating IV disc disorders
and is gradually replacing myelography and CT for the study of these
disorders (McCormick, 2000). MRI can demonstrate spinal cord or nerve root compression and indicate the degree of degenerative change within the IV disc (Fig. 4.37).
MRI is also an ideal screening procedure for the differential diagnosis
of structural disorders affecting the spinal cord and spinal nerve
roots.
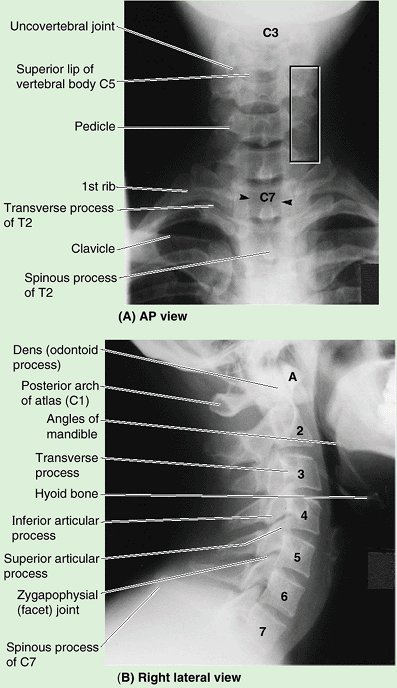 |
|
Figure 4.34. Radiographs of cervical region of vertebral column. A. The bifid spinous processes of C2–C6 vertebrae are overlapped by the radiotranslucency (black shadow) of the trachea. Arrowheads, margins of the column of air in the trachea; boxed area, the column of articular processes and the overlapping transverse processes. B. The anterior arch of the atlas (A)
lies in a plane that is anterior to the curved line joining the anterior borders of the vertebral bodies. The bodies of the C2–C7 vertebrae are numbered. Observe also the long spinous process of the vertebra prominens (C7). (Courtesy of Dr. J. Heslin, Toronto, Ontario, Canada.) |
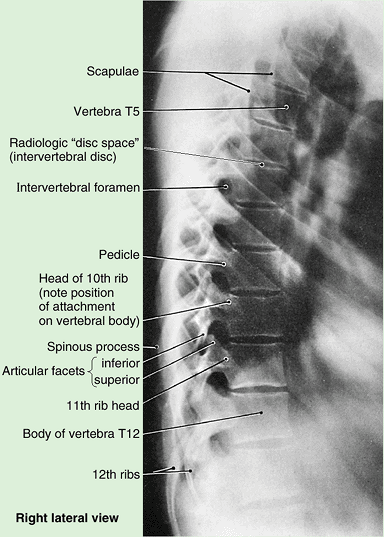 |
|
Figure 4.35. Radiograph of thoracic region of vertebral column.
In the lateral view of the thoracic vertebrae, the uniform vertebral bodies and radiographic disc “spaces” (caused by the radiolucency of the IV discs) are evident, except for the superior two or three, which are obscured by shoulder structures. The vertebral arch components, such as the transverse and spinous processes, are obscured by the posterior aspects of the overlapping ribs. |
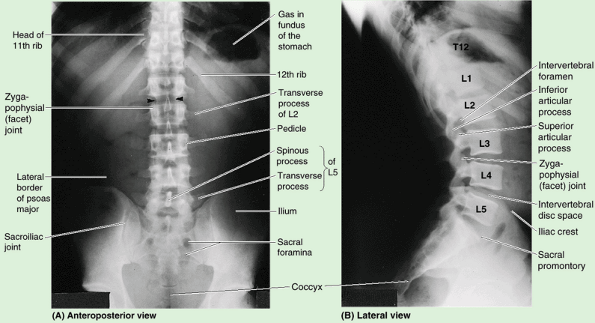 |
|
Figure 4.36. Radiographs of inferior thoracic and lumbosacral regions of vertebral column. A. Spinous processes in the midline overlap the vertebral canal (columnar shadow between arrowheads),
containing the spinal cord and spinal meninges (not visible). The coccyx is not well shown because of its oblique position relative to the film/detector. In addition, gas and feces in the rectum and sigmoid colon tend to obscure the coccyx. (Courtesy of Dr. J. Heslin, Toronto, Ontario, Canada.) B. The larger bodies of the lumbar vertebrae and the IV disc spaces, the IV foramina, and the angulation at the lumbosacral junction producing the sacral promontory are evident. The lumbar IV discs are wedge shaped, especially the L5–S1 disc. The zygapophysial (facet) joint between the superior articular process of vertebra L4 and the inferior articular process of vertebra L3 is clearly seen. |
 |
|
Figure 4.37. Effects of osteoporosis on vertebral column. A.
A loss of the horizontal trabeculae with thickening of the vertical trabeculae produces a pattern of vertical striation in the vertebral bodies, indicating early to moderate osteoporosis. B. Later, the striated pattern is lost as the continued loss of spongy bone produces uniform radiolucency (less white, more “transparent”). In contrast, the cortical bone of the vertebral endplates, while thinned, appears relatively prominent. C. Osteoporotic thoracic region of the vertebral column demonstrates excessive thoracic kyphosis, a result of the evident collapse of the vertebral bodies, which have become wedge-shaped (W), planar (P), and biconcave (B). |
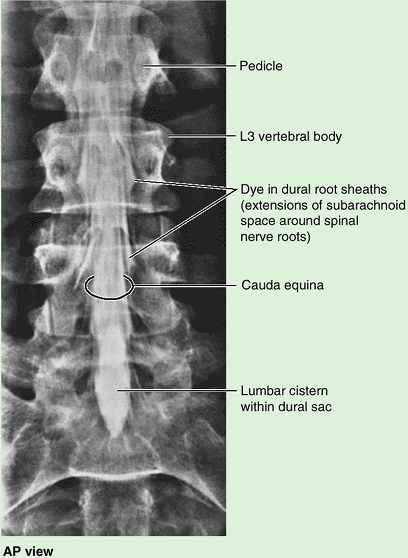 |
|
Figure 4.38. Myelogram of lumbar region of vertebral column.
Contrast medium was injected into the lumbar cistern. The lateral projections indicate the extensions of the subarachnoid space in the dural root sheaths around the spinal nerve roots. |
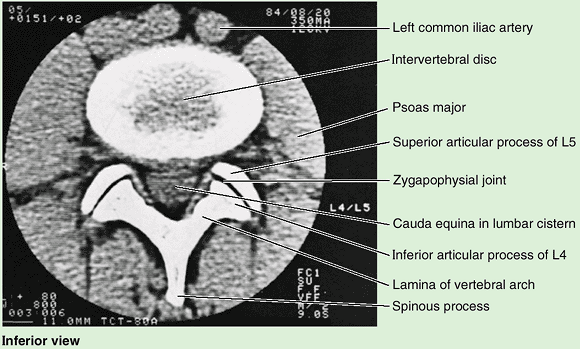 |
|
Figure 4.39. Transverse CT image of L4–L5 IV disc and vertebral arch.
The two parts of the IV disc (anulus fibrosis and nucleus pulposus) are apparent. The superior articular processes of the inferior vertebra (L5) “grip” the inferior articular processes of the superior vertebra (L4), disallowing rotatory movements but permitting flexion and extension. The cauda equina is seen running through the lumbar cistern within the vertebral canal; it is composed of all the spinal nerves inferior to the conus medullaris. |
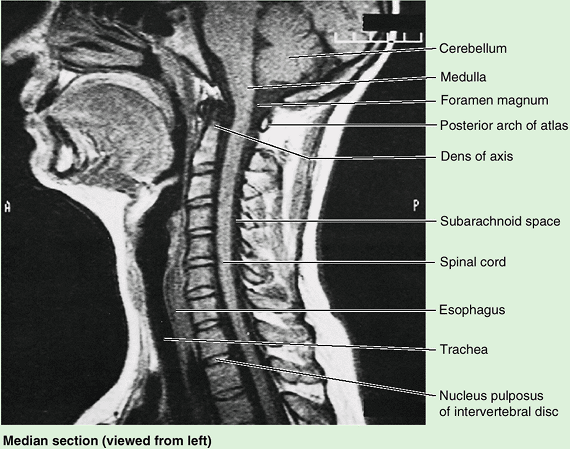 |
|
Figure 4.40. Midsagittal MRI of lower head and neck.
The cerebellum, medulla, spinal cord, and cervical region of the vertebral column are shown. The nuclei pulposi are evident in the IV discs of this young adult, interspersed between the spongy bone of the vertebral bodies. The anuli fibrosi and compact bone of the bodies produce little signal, and appear black. |
 |
|
Figure 4.41. MRI studies of vertebral column. A. T1-weighted sagittal MRI showing a C5–C6 IV disc herniation (arrow). The bright signal surrounding the spinal cord indicates the CSF. B. This axial (transverse) MRI demonstrates an IV disc herniation (arrow) in a patient with radiculopathy, a disorder of the spinal nerve roots (McCormick, 2000).
|
are often erroneously used as synonyms. Technically, however, in each
case the former is a gross anatomy term applied to parts of the adult
vertebrae, and the latter is an embryology term referring to parts of a
developing vertebra ossifying from primary centers. The vertebral body
includes the centrum and part of the neural arch; the vertebral arch is
thus less extensive than the neural arch, and the centrum is less
extensive than the vertebral body (O’Rahilly, 1986; Williams et al., 1995).
HA, Heilman AE, Stevens JM: Progressive myelopathy secondary to
odontoid fractures: Clinical, radiological, and surgical features. J Neurosurg 78: 579, 1993.